








La harpe urukéenne et le système auditif
le 8 janvier 2014 | par Philippe Roi et Tristan GirardPar Philippe Roi(1), Tristan Girard(2), Richard Dumbrill(3), Michel Leibovici(4)
Avec la participation de Paul Avan(5)
(1)Chercheur en Sciences Cognitives, spécialisé en Archéologie Cognitive ; (2)Chercheur en Sciences Cognitives ; (3)Professeur en Archéomusicologie, Directeur d’ICONEA, Institute of Musical Research, School of Advanced Study, University of London ; (4)Docteur en Biologie Cellulaire et Moléculaire de l’Université Paris VI, Chercheur au CNRS, Institut Cochin, INSERM U1016 ; (5)Directeur du Laboratoire de Biophysique Neurosensorielle de Clermont-FerrandU1016.
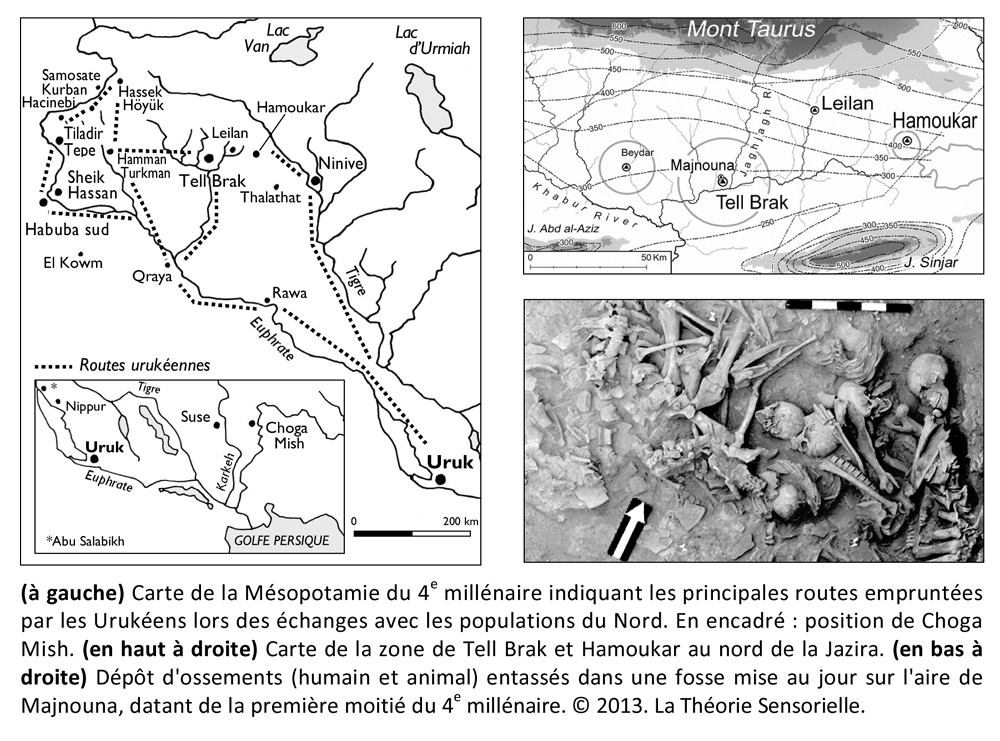
Afin d’obtenir toujours plus de produits qu’elles considéraient comme essentiels pour se distinguer du commun, les élites du sud de la Mésopotamie auraient multiplié les échanges avec celles du Nord, décuplant aussi les conflits comme semblent l’attester de récentes découvertes faites en Syrie sur les sites de Tell Brak, Majnouna et Hamoukar où plusieurs fosses communes ont été mises au jour. L’ensemble de ces fosses contenait des centaines, voire des milliers de squelettes de jeunes adultes, sur lesquels des marques laissent présumer une mort violente (1). Rappelons que Brak était à l’époque un carrefour commercial stratégique. Un sondage profond du site principal a permis de découvrir des éléments prouvant que les locaux importaient des matières premières du Levant Nord et d’Anatolie, qu’ils transformaient ensuite en biens manufacturés (2). Or, nous savons que si les terres de basse Mésopotamie, riches en limon, favorisaient les cultures et l’élevage, elles ne produisaient pas de pierres semi-précieuses, de métaux, de bois, de silex, ni de basalte ou d’obsidienne, et ne permettaient pas d’obtenir de denrées alimentaires comme l’huile d’olive, le vin ou le miel. En d’autres termes, le sol ne fournissait aucun produit susceptible d’épancher la frénésie ostentatoire des élites dont le pouvoir était en partie conditionné par ces signes extérieurs qui les différenciaient de leurs congénères et des cités voisines. Leurs seuls biens échangeables avec les villages du nord de la Mésopotamie consistaient donc en céréales et en ovins probablement dotés d’une toison d’une longueur et d’une finesse exceptionnelles.
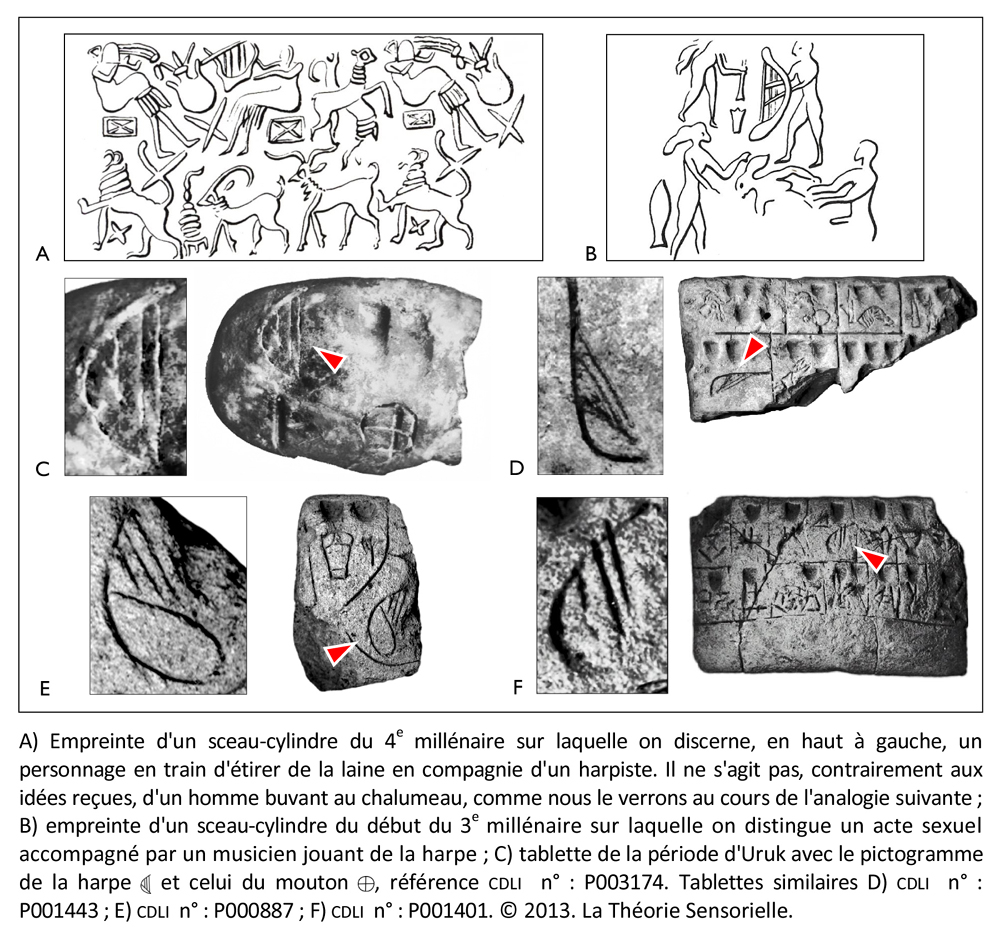
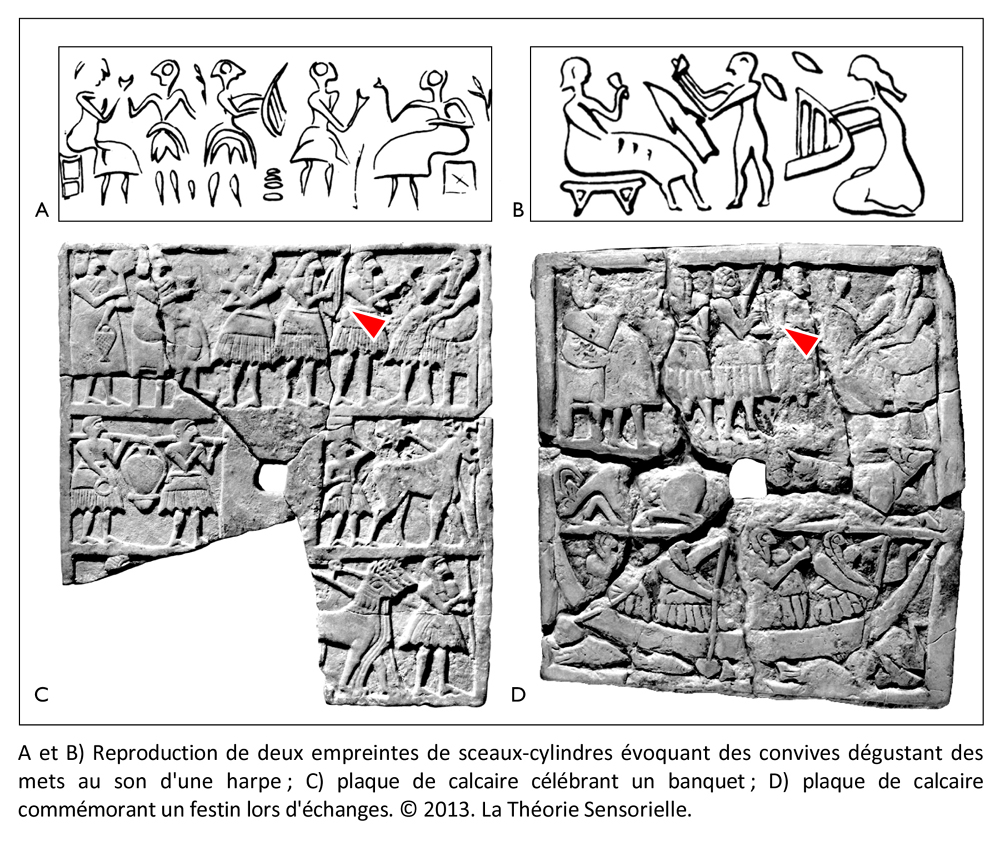
Contrairement aux éleveurs du Nord, les Urukéens pouvaient, en effet, se permettre de nourrir leurs animaux avec des céréales étant donné les quantités considérables d’orge et de blé qu’ils produisaient grâce à l’araire. Or, nous savons aujourd’hui que lorsque l’on nourrit des agneaux avec des céréales et du foin à forte teneur en sel, on obtient une laine des plus fines dont l’épaisseur peut ne pas dépasser 11,8 microns (3). Dès lors, on peut imaginer qu’entre le Nord et le Sud, les échanges consistaient en produits exotiques contre des céréales et des ovins dotés d’une toison hors du commun. Ces échanges ne devaient pas se faire sans de longues conversations dans des dialectes différents qui devaient se terminer au mieux par un festin et, dans le pire des cas, en conflits sanglants. L’histoire regorge d’exemples de différends commerciaux qui ont abouti en guerres. Il est donc possible qu’à la suite de massacres comme celui de Tell Brak, les protagonistes aient cherché un moyen de détendre l’atmosphère pour apaiser les tensions lors des négociations. Le plus subtil des arts, et aussi l’un des plus anciens, fut peut-être la solution trouvée par les Urukéens. En effet, c’est à cette époque qu’apparaît pour la première fois la harpe à trois ou quatre cordes, comme en témoigne une empreinte de sceau, datant de la moitié du 4e millénaire, retrouvée à Choga Mish à l’est d’Uruk en Elam (Iran moderne). Ce sceau représente un personnage assis tenant entre ses mains une harpe arquée à quatre cordes verticales tandis que deux autres individus manipulent respectivement un tambour en forme de bol et des claquoirs (4). Ainsi, les Urukéens auraient découvert dès le 4e millénaire, grâce à la harpe, que la musique avait la capacité de faire partager collectivement à des individus des sensations et des émotions que, dans d’autres circonstances, ils ne pouvaient éprouver que de façon individuelle. Cela signifie que la fonction première de la harpe et, par extension, de la musique, aurait été l’échange, car elle permettait d’emblée de trouver un terrain commun de compréhension, autrement dit de s’accorder. Dès lors, grâce à ce médiateur remarquable, la musique se serait insérée dans la démarche de communication entre les différentes populations, modifiant leur état d’esprit et participant au développement de leur empathie. En outre, les Urukéens auraient été les premiers à comprendre que, si la musique agissait de façon significative sur l’affectif de ses auditeurs, elle avait aussi une incidence sur leur comportement au cours des négociations. Or, nous savons aujourd’hui que si la musique ne modifie pas l’image d’un produit, elle agit de façon non-consciente sur l’humeur du consommateur qu’elle place dans des conditions plus favorables à l’achat. Dès lors, les aptitudes de la harpe à calmer les humeurs belliqueuses en engendrant des réactions émotionnelles l’auraient placée dans une position souveraine du lever au coucher du soleil. On la retrouve en effet dans les ateliers de poterie, les filatures, sur les barges qui transportent les marchandises, lors des banquets, voire pendant les rapports sexuels. Les Urukéens semblent si fiers de leur invention qu’ils la font graver sur des sceaux-cylindres afin que, même muette, la harpe résonne à l’oreille intérieure de ceux qui verront son empreinte sur les cachets des jarres. Ils la dessinent aussi sur des tablettes d’argile comme le suggèrent les mentions des termes sumériens ban.tur (ban signifiant « arc » et tur, « petit » donc « petit arc » = harpe) et le signe balag dont la phonétique est l’onomatopée « dub-dub », autrement dit un mot dont la sonorité rappelle ce qu’il désigne. Le pictogramme par lequel on le représente entre 3300 et 3000 avant notre ère évoque clairement une harpe à trois ou quatre cordes, mais cinq siècles plus tard, l’instrument deviendra un tambour (5).
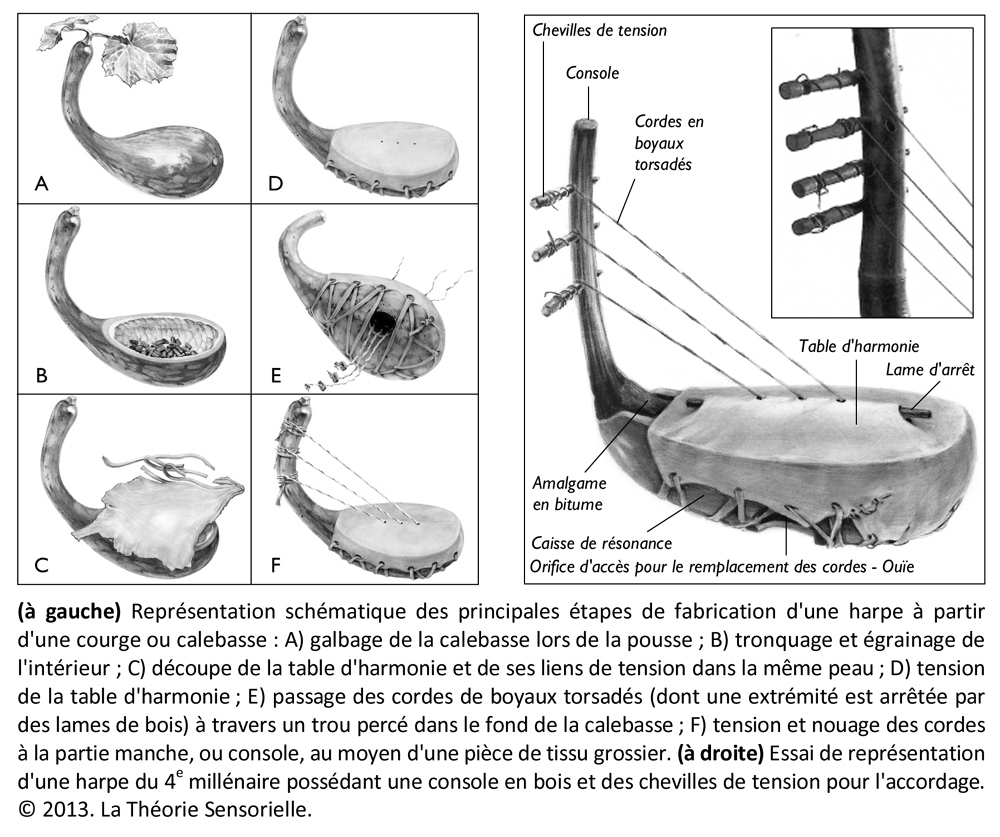
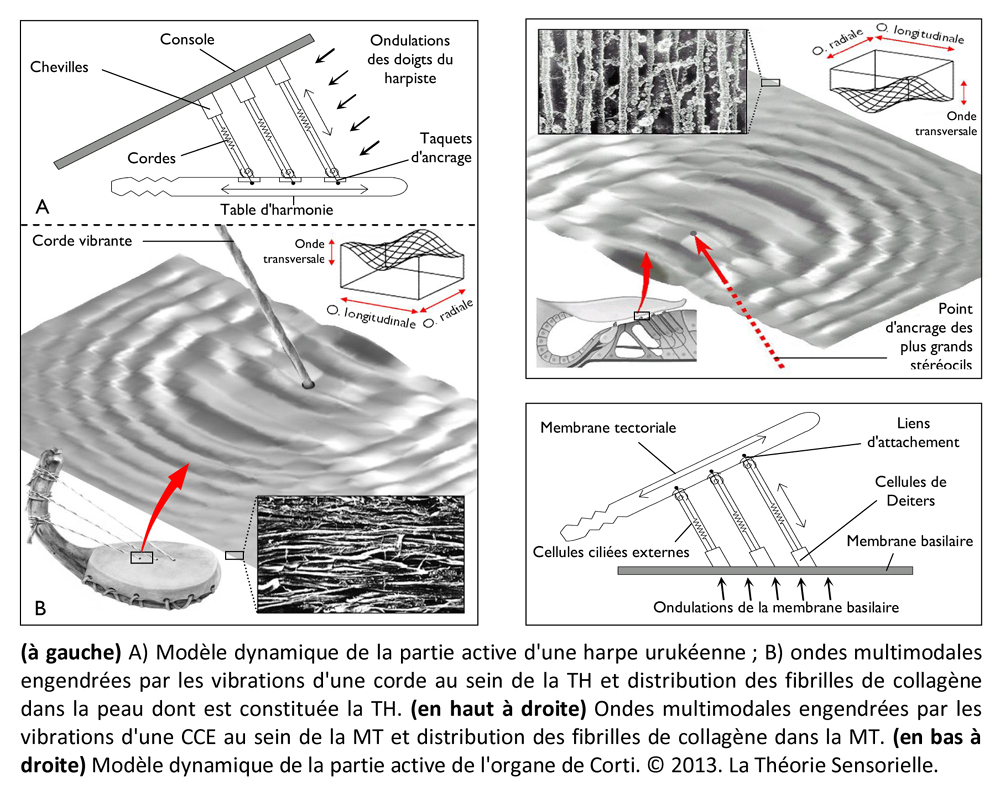
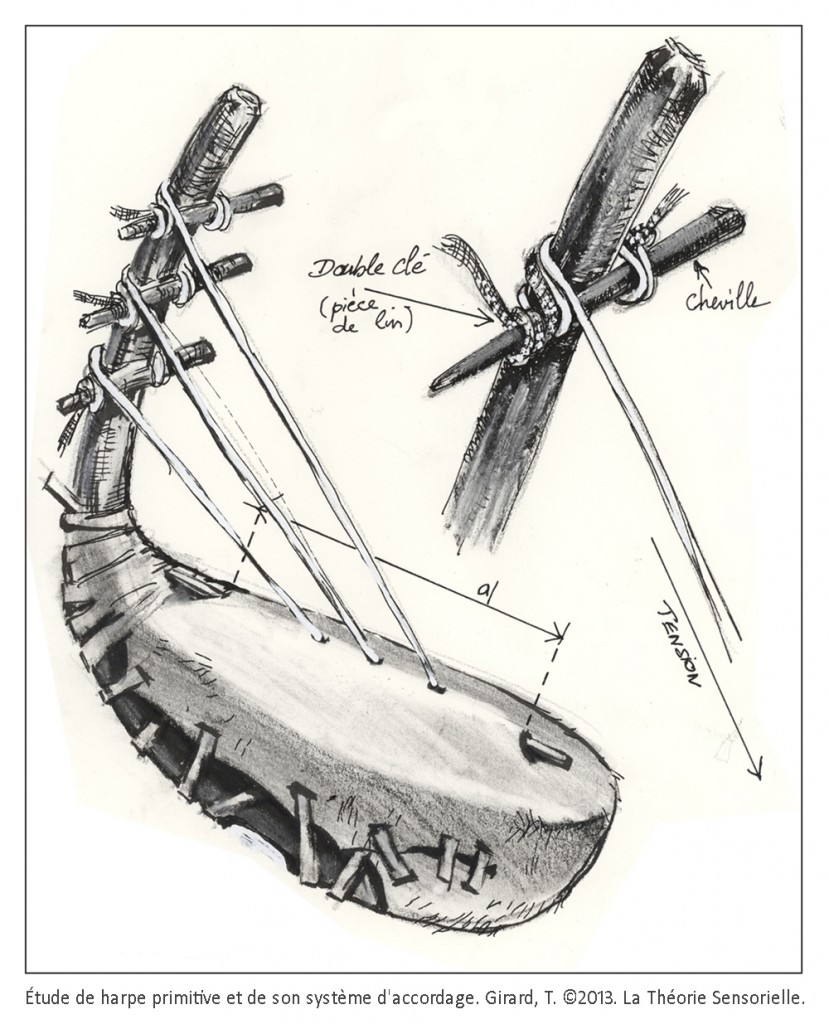
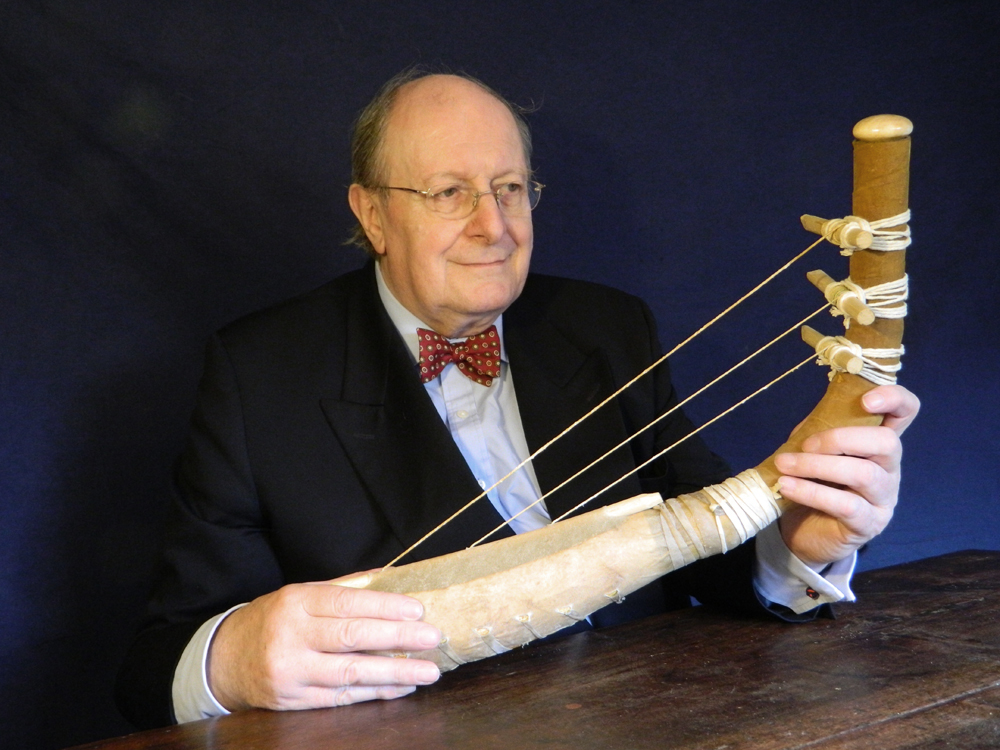
Toutes les harpes de la période d’Uruk/Djemdet Nasr avaient en commun d’être monoxyles ou tout au moins monostructurelles, en d’autres termes, leurs corps ne laissaient pas paraître de distinction morphologique entre la caisse de résonance et le manche qui deviendra console. Il semble que ces instruments étaient fabriqués à partir de cucurbitacées ou de bignoniacées – en général des courges ou calebasses – dont les formes naturelles étaient appropriées. Il est même possible que ces légumineuses ou ces fruits aient fait l’objet d’une domestication à l’aide de techniques simples pour prendre une forme s’accommodant à la fabrication de l’instrument souhaité (6). La partie évidée de la calebasse/gourde devait être ensuite recouverte d’une peau d’ovin, de bovin ou de porcin tendue en son dos, afin de former une caisse de résonance. Les cordes étaient faites à partir de boyaux torsadés ou de fibres végétales. Leur extrémité inférieure comportait un nœud d’arrêt. Elles étaient enfilées à travers une ou plusieurs baguettes percées, sous la table d’harmonie, afin que les nœuds ne s’échappent pas sous la tension. De même, les extrémités supérieures des cordes devaient être nouées à des bandes de tissu qui étaient enroulées autour de la console afin de maintenir leur tension pour assurer leur accordage. Au cours de cette période et surtout vers la fin du 4e millénaire, on observe un changement progressif de la caisse de résonance qui se développe en volume tandis que le manche/console se réduit. Il est probable que les caisses de résonance aient toujours été faites à partir de calebasses ou de cucurbitacées, mais que les consoles aient été fabriquées en bois pour être plus résistantes aux pressions des chevilles dont les Urukéens se seraient servis plus tard pour assurer la tension des cordes. Les harpes du 4e millénaireétaient toutes probablement de petites tailles. Elles ne devaient avoir en général que trois cordes tendues sur un plan d’inclinaison d’environ 110 degrés, ce qui suggère une disposition anhémitonique s’étalant sur une quinte musicale contenant peut-être une tierce. Ces harpes sont toujours représentées en milieu rural à en juger par la présence d’ovins et autres ruminants à leurs côtés, ce qui signifie qu’elles n’étaient pas destinées à des rituels religieux, mais avaient au 4e millénaire, une utilité pratique, comme le confirment les empreintes de sceaux-cylindres. Cependant, dès le troisième millénaire, elles sont toujours représentées en présence d’Inanna, divinité protectrice de la cité d’Uruk, quand elles ne symbolisent pas la déesse elle-même. Il existe des textes où il est fait mention des animaux d’Inanna et de ses attributs parmi lesquels figurent le roseau, la palme, son astre Vénus et la harpe.
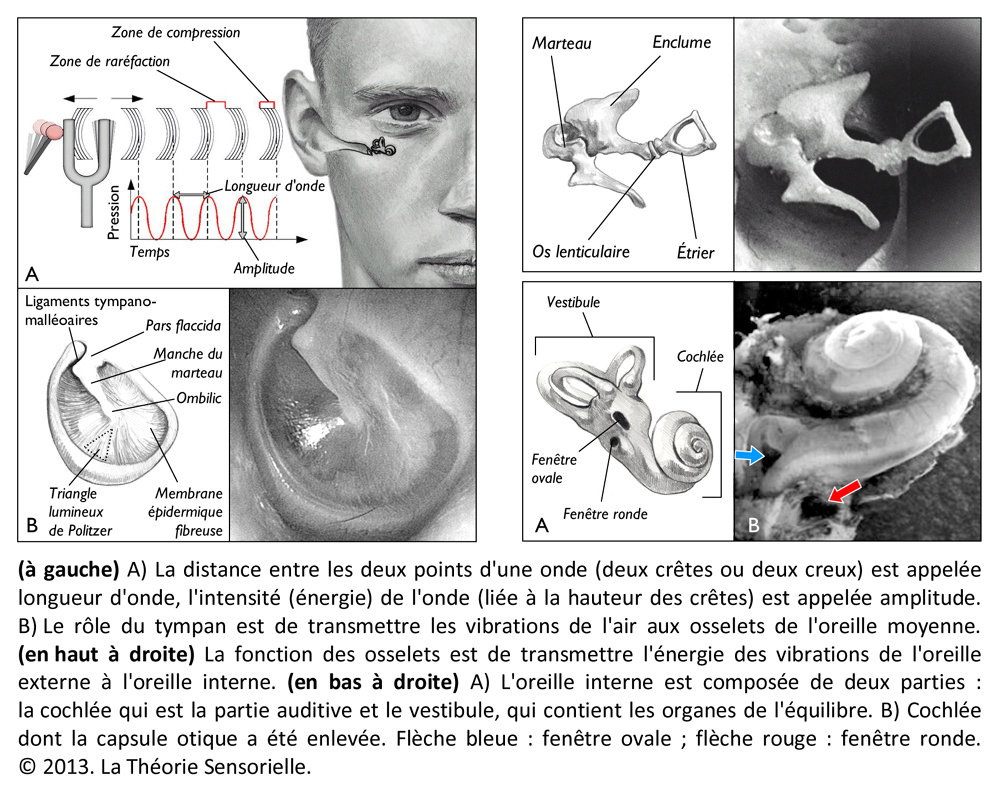
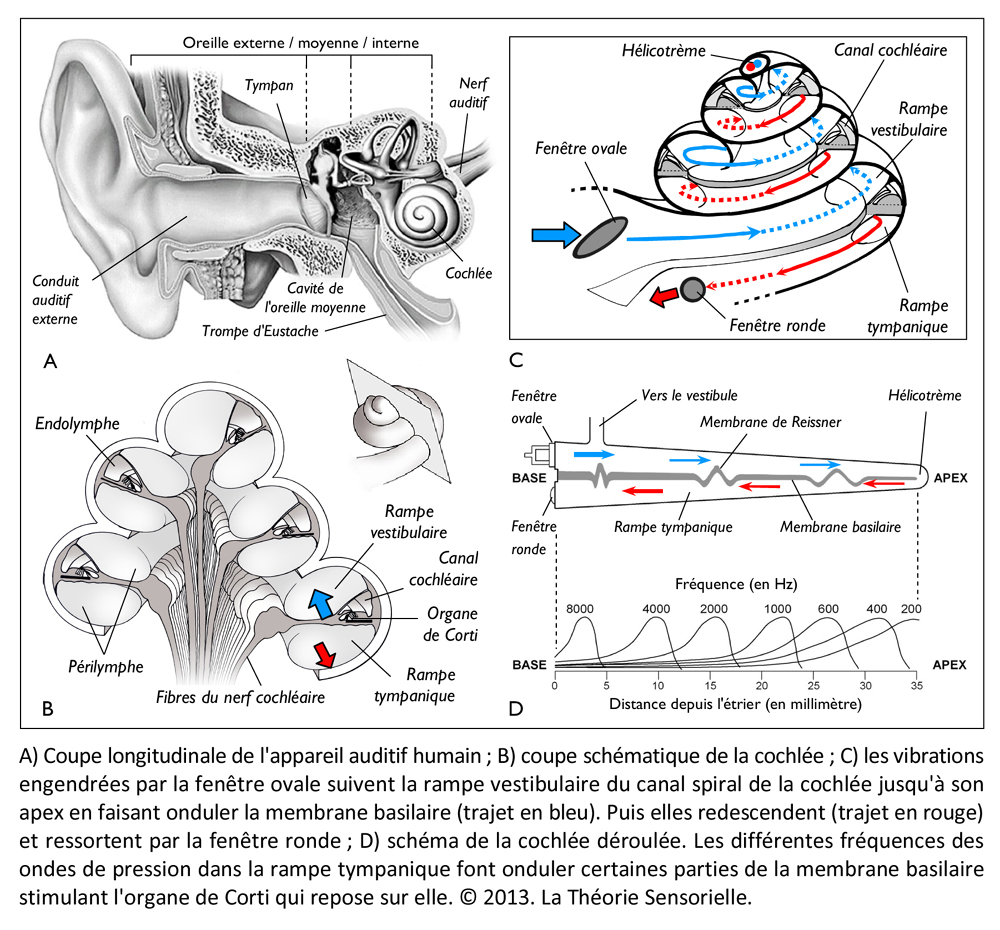
La harpe étant décrite et replacée dans son contexte d’origine, il est intéressant de constater que sa conception repose sur les principes fondamentaux de l’audition. Chez l’homme, la perception du son est tout à fait remarquable. Ce que nous appelons des sons n’est en fait que des mouvements de l’air, de simples variations périodiques de pression qui se propagent comme un front d’onde à la vitesse de trois cents mètres par seconde. Lorsqu’elles parviennent à l’oreille humaine, elles sont canalisées par le conduit auditif jusqu’à une fine membrane appelée tympan qui sépare l’oreille externe de l’oreille moyenne. Les vibrations du tympan, sous l’effet des variations de pression acoustique, sont transmises à une chaîne de quatre osselets (marteau, enclume, os lenticulaire et étrier) contenus dans la partie pétreuse de l’os temporal. Ces osselets, maintenus entre eux par des articulations et des ligaments, permettent aux vibrations de passer, sans perte d’énergie, du milieu aérien au milieu aqueux de l’oreille interne. Cette dernière, située dans l’os temporal, est une structure complexe constituée d’un labyrinthe osseux et de plusieurs cavités remplies de liquide. Ce réseau continu, composé de canaux, d’ampoules et d’un limaçon enroulé comme une coquille d’escargot, abrite en fait deux organes sensoriels bien distincts. D’une part, le système vestibulaire spécialisé dans la détection et l’adaptation des mouvements du corps dans l’espace et, d’autre part, la cochlée ou limaçon osseux qui constitue l’organe récepteur de l’audition.
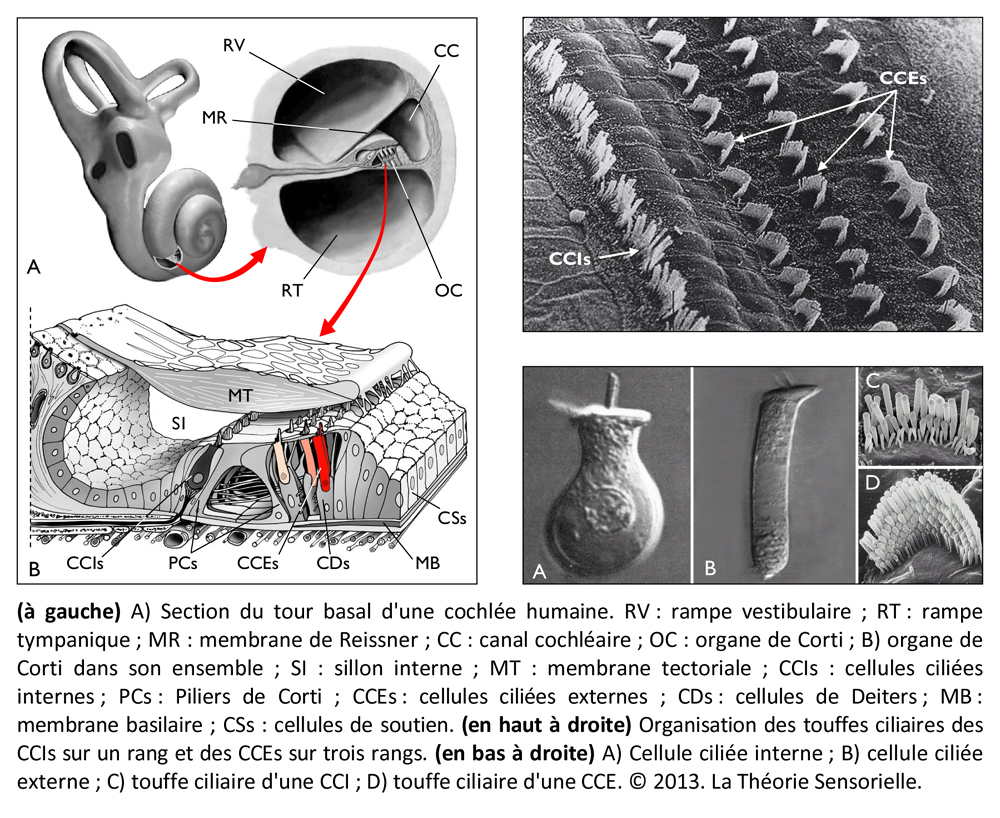
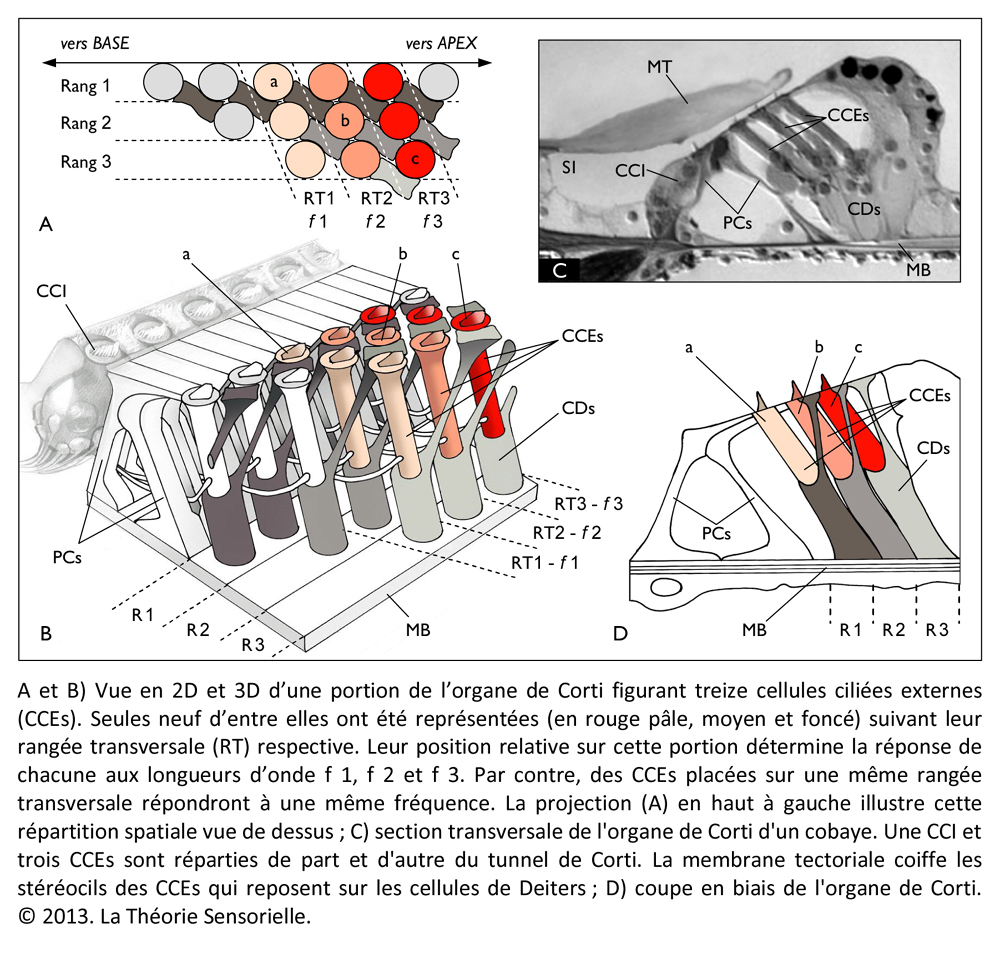
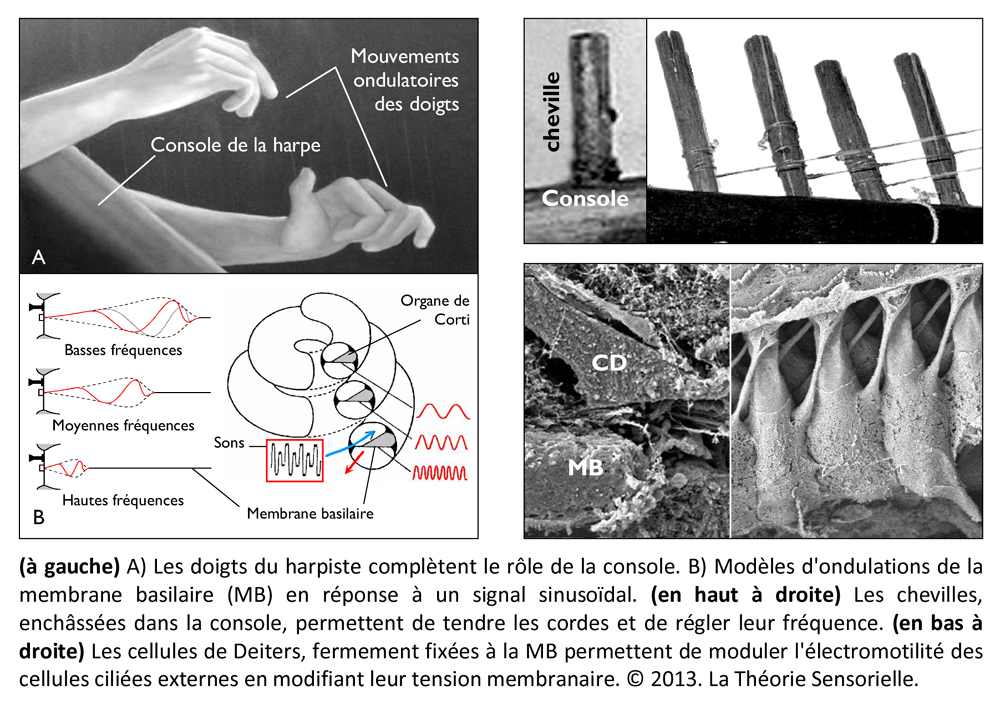
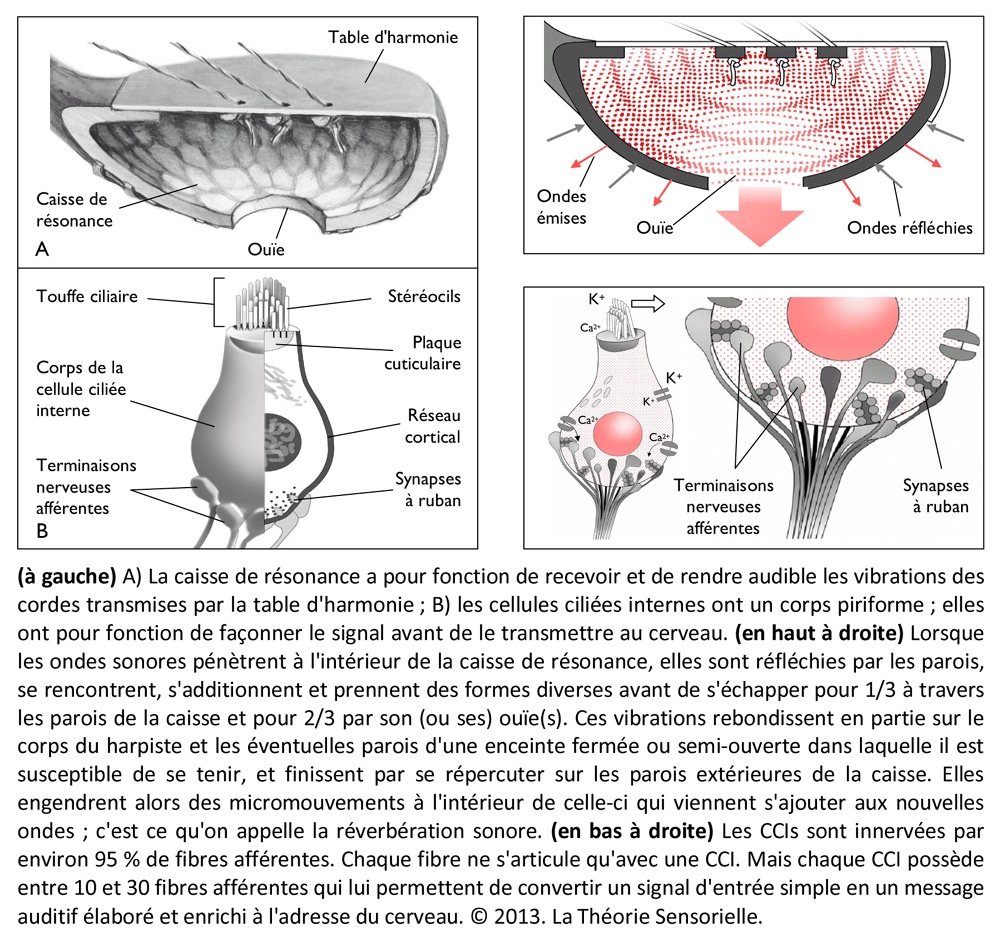
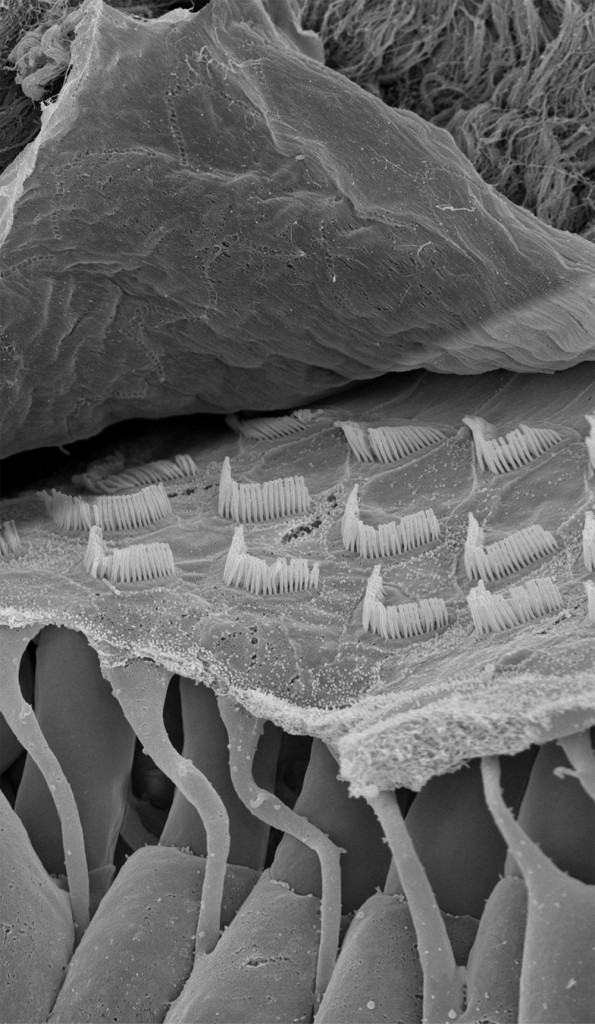
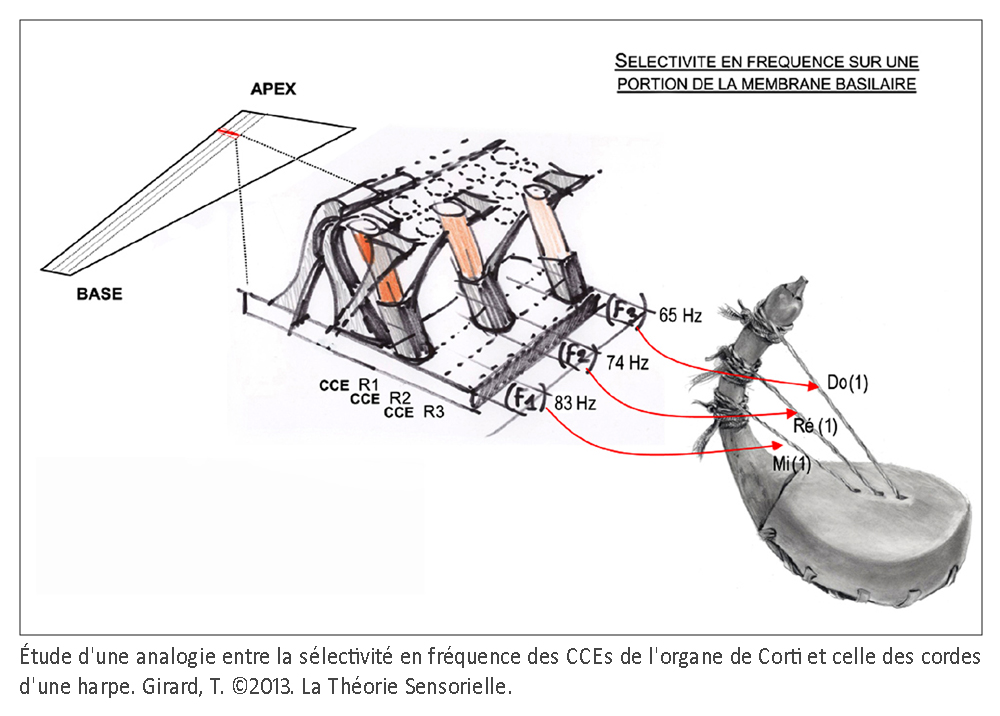
La cochlée (du latin cochlea qui signifie « escargot ») est une petite structure spiralée de 1,2 mm de diamètre pour 35 mm de long qui prend naissance dans la partie antérieure du vestibule et décrit deux tours trois quarts autour d’un pilier osseux appelé columelle. À son extrémité basale se trouvent la fenêtre ovale et la fenêtre ronde, qui séparent l’oreille moyenne de l’oreille interne. La cochlée est divisée sur sa longueur en trois compartiments : de part et d’autre se trouvent deux compartiments, la rampe vestibulaire et la rampe tympanique, remplis d’un même liquide, la périlymphe ; au milieu se situe le canal cochléaire limité à sa partie inférieure par la membrane basilaire et à sa partie supérieure par la membrane de Reissner. Le canal cochléaire abrite une structure complexe appelée organe de Corti qui repose sur la membrane basilaire et s’étend sur toute la longueur de la cochlée. C’est cet organe qui assure la transformation de l’énergie vibratoire en signal électrique interprétable par le cerveau. Il est constitué de 15 000 cellules ciliées externes qui se répartissent sur trois ou quatre rangées et de 3500 cellules ciliées internes qui s’alignent selon une seule rangée. Les corps cellulaires de celles-ci baignent dans la périlymphe tandis que leurs touffes ciliaires se dressent dans le canal cochléaire rempli d’endolymphe. L’étanchéité ionique du canal cochléaire est assurée à sa base par la lame réticulaire, qui résulte de l’association très étroite entre les apex des cellules sensorielles et des cellules de soutien de l’organe de Corti, et au sommet, par la membrane de Reissner qui forme le « toit » du canal cochléaire. Lorsque l’étrier se déplace sous la poussée des vibrations du tympan, il met en mouvement le liquide incompressible contenu dans la rampe vestibulaire et le fait remonter jusqu’au sommet de la cochlée où il contourne l’hélicotrème avant de redescendre par la rampe tympanique, mettant finalement en mouvement la fenêtre ronde qui se situe à l’autre extrémité du système. Les mouvements du liquide font onduler la membrane basilaire dont chaque portion vibre à une fréquence donnée. Ces vibrations entraînent un déplacement des cellules ciliées externes, qui sont enchâssées entre la membrane basilaire (par leur pôle basal via les cellules de Deiters) et la membrane tectoriale (via leur touffe ciliaire). Il s’ensuit un mouvement relatif de cisaillement des touffes ciliaires des cellules ciliées externes vis-à-vis de la membrane tectoriale qui provoque la déflexion des stéréocils. Cette dernière conduit à la dépolarisation cellulaire par ouverture mécanique de canaux ioniques. La dépolarisation engendre une contraction rapide des cellules ciliées externes ou électromotilité qui, à son tour, augmente l’amplitude des vibrations de la membrane basilaire. Ce phénomène d’amplification non linéaire a pour conséquence d’augmenter les stimuli faiblement intenses qui pourraient ne pas être perçus sans affecter les stimuli de forte intensité dont l’amplification endommagerait les cellules ciliées. Sous l’effet de cette amplification, les stéréocils des cellules ciliées internes, véritables récepteurs sensoriels de l’organe de l’audition, sont entraînés par la membrane tectoriale et défléchis à leur tour. La dépolarisation de la cellule ciliée interne engendre alors la libération d’un neurotransmetteur à son pôle basal, le glutamate, qui donne naissance à un train de potentiels d’action acheminés via le nerf auditif jusqu’au cerveau.
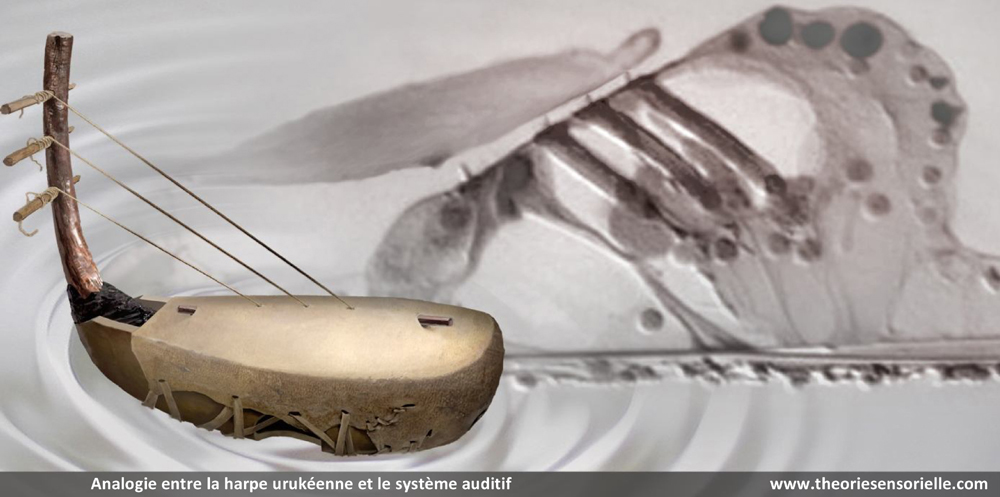
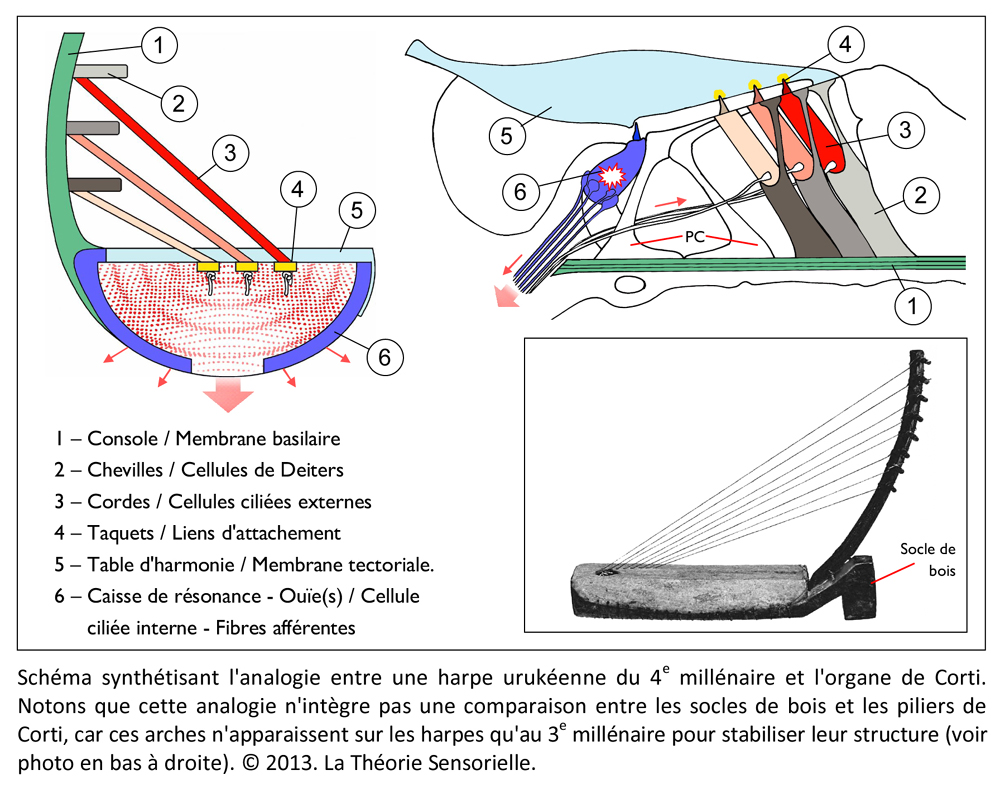
À ce stade, une première comparaison peut être faite entre le manche de la harpe (ou console) et la membrane basilaire (MB). La console, à laquelle sont fixées les chevilles autour desquelles sont attachées l’une des extrémités des cordes, peut en effet être comparée à la membrane basilaire à laquelle sont fixées les cellules de Deiters supportant elles-mêmes l’une des extrémités des cellules ciliées externes (CCEs). La console est un élément primordial, car c’est sur celle-ci que reposent directement ou indirectement tous les éléments qui constituent une harpe. L’importance fonctionnelle de la membrane basilaire est elle aussi primordiale, car c’est elle qui supporte directement ou indirectement les différents éléments qui constituent l’organe de Corti. Les doigts du harpiste font vibrer les cordes avec plus ou moins d’intensité, complétant ainsi la fonction de la console qu’il maintient avec une paume contre sa poitrine. De façon identique, les vibrations sonores transmises par la périlymphe provoquent des mouvements vers le haut et vers le bas de la membrane basilaire en une position précise qui font vibrer avec plus ou moins d’intensité les cellules ciliées externes. À l’instar de la console, la membrane basilaire est immobile en tout autre endroit le long de l’organe de Corti.
Une deuxième comparaison peut être faite entre les chevilles insérées dans l’épaisseur de la console et les cellules de Deiters (CDs) dont la base est fermement fixée sur la membrane basilaire. Les chevilles ont pour fonction de maintenir solidement les cordes tendues entre la console et la table d’harmonie. De façon analogue, les cellules de Deiters servent à maintenir fermement les cellules ciliées externes (CCEs) tendues entre la membrane basilaire (MB) et la membrane tectoriale (MT). De la même façon que la rotation des chevilles dans un sens ou dans l’autre permet au musicien de tendre ou de détendre les cordes pour les accorder afin de les faire vibrer à la fréquence désirée, il a été récemment démontré que la déformation des cellules de Deiters pouvait moduler la vibration des CCEs auxquelles elles sont associées en modifiant leur tension membranaire (7). Cette modification locale doit contribuer à affiner la réponse au stimulus acoustique.
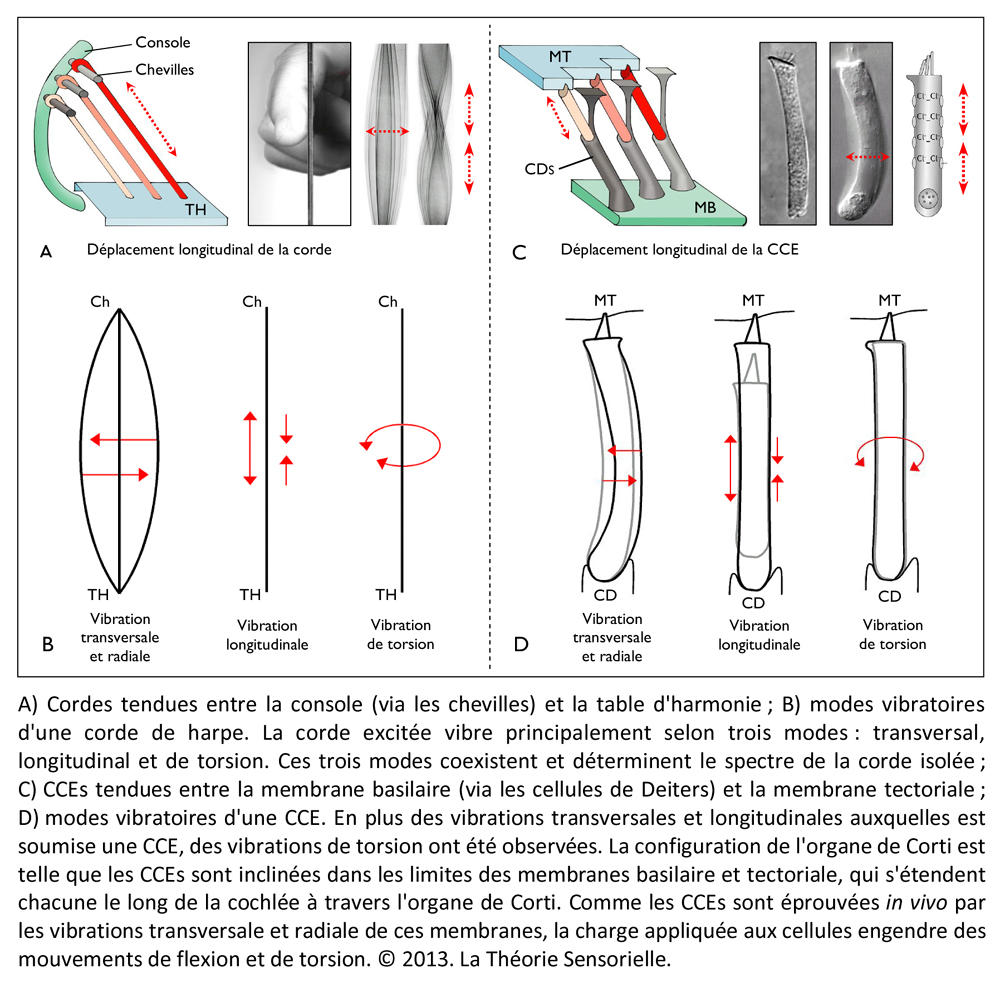
Une troisième comparaison peut être faite entre les cordes de la harpe et les cellules ciliées externes de la cochlée. Les cordes sont fixées par une extrémité aux chevilles et par l’autre à la table d’harmonie. Cette attache à ces deux structures confère aux cordes un degré de tension suffisant pour vibrer lorsqu’elles sont pincées. Le pincement d’une corde entraîne un mouvement radial de celle-ci en même temps qu’un léger pivotement de la corde sur elle-même du fait qu’elle est torsadée. Il s’ensuit une propagation ondulatoire dans le sens longitudinal depuis leur point d’ancrage jusqu’au sein de la table d’harmonie. D’une façon identique, les cellules ciliées externes (CCEs) sont fixées par leur extrémité basale aux cellules de Deiters, et par leur touffe ciliaire à la membrane tectoriale. Ainsi maintenues, les CCEs présentent des mouvements très rapides de contraction/décontraction de leur corps cellulaire – appelés électromotilité cellulaire – couplés à des oscillations périodiques de leur touffe ciliaire. Associés à l’électromotilité, des pivotements de la base des CCEs ont été décrits conduisant à un déplacement radial de la cellule. Il convient d’ajouter que la harpe ne connaît que le mode de mise en vibration des cordes par pincement. Leur disposition ne permet pas en effet de raccourcir l’espace de vibration par appui sur le manche, comme c’est le cas avec un violon. Chaque corde résonne donc à vide et ne produit qu’un seul son. De même, chaque rangée transversale de cellules ciliées externes, de par sa position le long de l’organe de Corti et ses caractéristiques structurales, ne répond qu’à une seule fréquence de stimulation. Enfin, à l’instar des doigts du harpiste qui peuvent bloquer ou étouffer les vibrations de certaines cordes et laisser les autres vibrer librement – voire par sympathie – de nombreuses fibres efférentes se projettent depuis le système nerveux central sur les CCEs par l’intermédiaire de synapses cholinergiques afin de moduler l’intensité de la réponse de ces dernières (8).
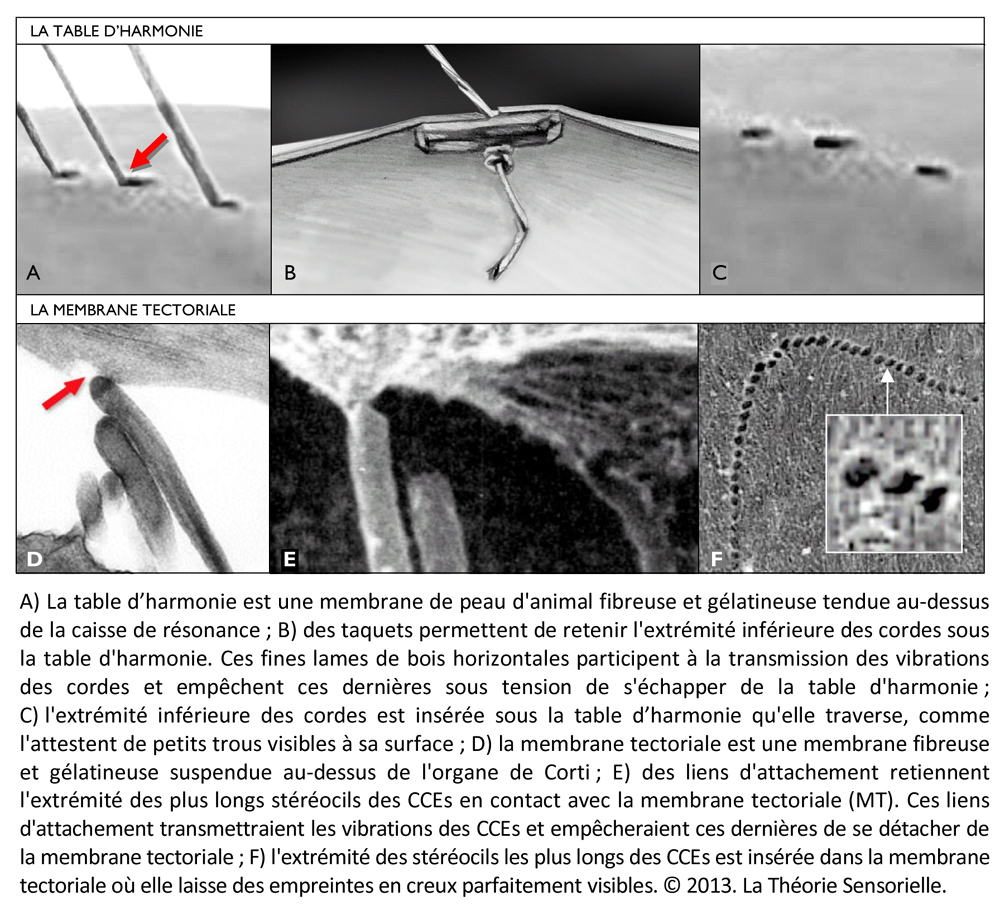
Une quatrième comparaison peut être faite entre la table d’harmonie et la membrane tectoriale. La table d’harmonie est une membrane de peau d’animal fibreuse et gélatineuse tendue au-dessus de la caisse de résonance. L’une des extrémités des cordes est insérée sous la table d’harmonie qu’elle traverse, comme l’atteste la présence de petits trous visibles lorsqu’on les retire. La surface des cordes étant très restreinte, leurs vibrations ne produisent que des sons très faibles. En revanche, si les vibrations des cordes sont transmises à une surface plus grande possédant des propriétés conductrices, les sons qu’elles produisent durent moins longtemps, mais sont d’une plus grande intensité. Outre l’amplification du signal provenant des cordes, la table d’harmonie assure sa transmission vers la caisse de résonance. De même, la membrane tectoriale est une membrane fibreuse et gélatineuse qui recouvre l’organe de Corti. Les extrémités des cils les plus longs des cellules ciliées externes sont insérées dans cette membrane à la surface de laquelle elles laissent des empreintes en creux parfaitement visibles au microscope électronique. Lorsque les CCEs vibrent, leurs touffes ciliaires s’inclinent et amplifient alors le mouvement relatif entre la membrane basilaire et la membrane tectoriale. Le signal amplifié est alors transmis aux cellules ciliées internes. Ainsi, la membrane tectoriale, tout comme la table d’harmonie, joue un rôle primordial dans la transmission de la stimulation sonore.
Une cinquième comparaison peut être faite aussi entre les taquets d’ancrage et les liens d’attachement de la membrane tectoriale. Les taquets d’ancrage sont de fines lames de bois fixées horizontalement sous la table d’harmonie. Ils participent à la transmission des vibrations et empêchent que les nœuds aux extrémités inférieures des cordes ne s’échappent sous la tension. D’une façon identique, des liens d’attachement, insérés sous l’épaisseur de la membrane tectoriale, ont été observés. Leur présence a pour but d’arrimer l’apex de la plus haute rangée de stéréocils des touffes ciliaires des cellules ciliées externes dans la membrane tectoriale. Cet ancrage promeut l’efficacité de la vibration des CCEs et, dans une moindre mesure, les protège de trop fortes tensions (9).
Enfin, une sixième comparaison peut également être faite entre la caisse de résonance (et son ouïe) et la cellule ciliée interne (et ses fibres afférentes). En organologie, on appelle caisse de résonance la partie d’une harpe qui a pour rôle de recevoir et de rendre audible les vibrations produites par les cordes et la table d’harmonie. Lorsque les ondes sonores pénètrent à l’intérieur de la caisse, elles se heurtent à ses parois. Une partie des ondes est absorbée tandis que l’autre rebondit. Les ondes qui se réverbèrent se rencontrent et s’additionnent et il en résulte un son plus perceptible, qui s’échappe notamment par une ou des ouvertures, situées à la base ou sur les côtés de la caisse, appelées les ouïes. Aux vibrations en deux dimensions, générées par les cordes et transmises par la table d’harmonie, la caisse de résonance apporte au son amplifié une troisième dimension acoustique : sa forme. En effet, la configuration de la caisse façonne naturellement le son, qui peut même être modulé grâce à des pièces de bois rectangulaires, appelées barrages, fixées sur sa base intérieure et sous la table d’harmonie pour guider les ondes et l’enrichir de nouvelles euphonies. Ainsi, selon la complexité et la qualité des barrages, le son et les instruments peuvent respectivement prendre des formes élaborées et différer en puissance et en sonorité. La cellule ciliée interne (CCI) est, quant à elle, la véritable cellule sensorielle de l’organe de Corti dans la mesure où elle perçoit la stimulation extérieure et la transmet au cerveau. La CCI est stimulée mécaniquement par les vibrations transmises par la membrane tectoriale qui défléchissent sa touffe ciliaire apicale. Cette déflexion entraîne une entrée de calcium qui génère à son tour, à la base de la cellule, une libération de neurotransmetteur qui est corrélée à la stimulation. Il s’ensuit dans les fibres nerveuses afférentes des potentiels d’action qui sont conduits par le nerf auditif aux centres nerveux supérieurs. Il a été démontré que les constituants pré et postsynaptiques présentent une distribution morpho-fonctionnelle qui permet à la CCI de façonner sa réponse électrique (10). Ainsi, à l’instar de la caisse de résonance et des ouïes, la CCI et les fibres nerveuses afférentes convertissent un signal d’entrée simple – oscillation de la touffe ciliaire – en un signal de sortie élaboré.
NOTES
(1) McMahon, A. (2007) Soltysiak, A. (2007).
(2) Oates, J. (2007).
(3) Lupton, C.J. et al. (2007).
(4) Delougaz, P.P.; Kantor, H.J. (1972).
(5) Dumbrill, R. (1998). Période au cours de laquelle les cités-États sont en guerre.
(6) Dumbrill, R. (2012).
(7) Yu, N.; Zhao, H.-B. (2009).
(8) Frolenkov, G.I. (2006) Maison, S.F. et al. (2007) Richard, C. (2010).
(9) Richardson, G.P. et al. (2008) Verpy, E.; Leibovici, M. et al. (2011).
(10) Safieddine, S.; El-Amraoui, A.; Petit, C. (2012).
APPENDICE
Il n’est guère envisageable que les Mésopotamiens de la culture d’Uruk aient inventé la harpe au 4e millénaire sans étudier les sons produits par deux cordes vibrantes, faites de la même matière et tendues de la même façon, mais de longueurs différentes. Ils auraient alors constaté que si la corde était longue, il y avait peu de vibrations et le son qu’elle produisait était grave, et que si la corde était courte, les vibrations étaient plus nombreuses et le son généré était aigu. C’est ce que les physiciens appelleront six mille ans plus tard la fréquence. Il est probable que les Urukéens aient aussi constaté qu’une corde plus épaisse faite de boyau torsadé produisait un son plus grave qu’une corde mince, également faite de boyau torsadé et que, en augmentant la tension de la corde, le son devenait plus aigu. C’est en variant finement la tension des cordes qu’ils apprirent vraisemblablement à accorder leur harpe comme l’atteste la présence de chevilles fixées au sommet de la console de leurs instruments. Au cours de ces expériences, on peut supposer qu’ils constatèrent que l’association de sons était agréable à l’ouïe uniquement dans deux cas. Tout d’abord, lorsque certains rapports de longueurs de cordes permettaient d’obtenir des sons qui s’accordaient entre eux. Ensuite lorsque le rapport des longueurs des cordes équivalait à 2/1; 3/2; 4/3; 5/4 et 6/5, ce qui permettait d’obtenir une consonance parfaite.
BIBLIOGRAPHIE DE LA HARPE
reni, C.S.; Kim, D., ‘The influence of background music on shopping behavior: Classical versus top-forty music in a wine store.’ Advances in Consumer Research. Vol. 20. Published by the Academic Resource Center (1993) pp. 336-340.
Biggs, R.D., ‘The Sumerian Harp.’ The American Harp Journal. Vol. 1/3. Published by The American Harp Society (1968) pp. 6-12.
Caubet, A., ‘La Musique à Ougarit : nouveaux témoignages matériels.’ Ugarit, Religion and Culture: Proceedings of the International Colloquium on Ugarit, Religion and Culture, Edinburgh, July 1994. Essays presented in honour of Professor John C.L. Gibson. N. Wyatt ; W. G.E. Watsonand ; J.B. Lloyd (ed.) (1996) pp. 9-31.
Chadefaux, D.; Le Carrou, J.-L.; Fabre, B.; Daudet, L.; Quartier, L., ‘Experimental Study of the Plucking of the Concert Harp.’ Proceedings of 20th International Symposium on Music Acoustics (Associated Meeting of the International Congress on Acoustics) 25-31 August, Sydney and Katoomba (2010) pp. 1-5.
Chadefaux, D.; Le Carrou, J.-L.; Buys, K.; Fabre, B.; Daudet, L., ‘Étude expérimentale du pincement d’une corde de harpe.’ Actes du 10ème Congrès Français d’Acoustique, Lyon, 12-16 Avril 2010. Publié en ligne (2010).
Chadefaux, D.; Le Carrou, J.-L.; Fabre, B.; Daudet, L., ‘Experimentally based description of harp plucking.’ Journal of Acoustical Society of America. Vol. 131/1. Published by the Acoustical Society of America, Melville (2012) pp. 844-855.
Collon, D., ‘La musique dans l’art mésopotamien.’ Dossiers d’Archéologie. N° 310. Éditions Faton (2006) pp. 6-15.
Delalleau, A.; Josse, G.; Lagarde, J.M., ‘Un modèle hyperélastique à réorientation de fibres pour l’analyse des caractéristiques mécaniques de la peau.’ 9ème Colloque Nationale en Calcul des Structures, Giens 2009. Communication publiée en ligne.
Delougaz, P.; Kantor, H., ‘Choga Mish – Volume I – The First Five Seasons of Excavations, 1961-1971.’ The University of Chicago – Oriental Institute Publications. A. Alizadeh (ed.) Vol. 101. Published by the Oriental Institute of Chicago (1996) Plate 45-N.
Delougaz, P.P.; Kantor, H.J., ‘New Evidence for the Prehistoric & Protoliterate culture development of Khuzestan.’ The Memorial Volume of the Vth International Congress of Iranian Art & Archaeology. Tehran – Isfahan – Shiraz. 11th – 18th April 1968. Vol. 1. Special Publication of the Ministry of Culture and Arts, Tehran (1972) pp. 14-33.
Dubé, L.; Chebat, J.-C.; Morin, S., ‘The effects of background music on consumers desire to affiliate in buyer-seller interactions.’ Psychology and Marketing. Vol. 12/4. Published by Wiley-Blackwell (1995) pp. 305-319.
Dumbrill, R., ‘Appendix.’ A Queen’s Orchestra at the Court of Mari: New Perspectives on the Archaic Instrumentarium in the Third Millenium. M. Marcetteau. ICONEA Proceedings 2008 (2008) pp. 73-75.
Dumbrill, R., ‘Harps.’ The Archaeomusicology of the Ancient Near East. Trafford Publishing (2005) pp. 179-226.
Dumbrill, R., Götterzahlen and Scale Structure (1997).
Dumbrill, R., ‘Music Theorism in Ancient World.’ ICONEA Proceeding 2009-2010 (2010) pp. 107-132.
Erman, A.; Ranke, H., ‘Les divertissements.’ La civilisation égyptienne. Éditions Payot (1985) p. 318.
Farmer, H.G., ‘The Music of Ancient Mesopotamia.’ New Oxford History of Music. Ancient and Oriental Music. Vol. 1. Published by Oxford University Press (1957) pp. 228-254.
Frankfort, H., Kingship and the Gods, a Study of Ancient Near Eastern religion as the Integration of Society and Nature. Published by the University of Chicago Press (1948).
Galpin, F.W., ‘The Sumerian Harp of Ur.’ Music & Letters. Vol. 10/2. Published by the Oxford University Press (1929) pp. 108-123.
Glattauer, A., À l’origine de la harpe. Éditions Buchet-Chastel (1999) pp. 13-23.
Guéguen, N.; Jacob, C.; Lourel, M.; Le Guellec, H., ‘Effect of Background Music on Consumer’s Behavior: A Field. Experiment in a Open-Air Market.’ European Journal of Scientific Research. Vol. 16/2. Published by the Scientific Research Platform – SRP (2007) pp. 268-272.
Hartmann, H., Die Musik der Sumerischen Kultur. Frankfurt am Main (1960).
Helmholtz, H.L.F., On the sensations of tone, as a physiological basis for the Theory of Music. Third Edition. Published by Longmans, Green and Co. (1865) reprinted by permission of Dover Publication, Inc. (1954).
Ivanov, V.V., ‘An Ancient Name of the Lyre.’ Archiv Orientální. Vol. 67. Published by the Oriental Institute, Academy of Sciences of the Czech Republic (1999) pp. 585-600.
Joannès, F., ‘Musique.’ Dictionnaire de la civilisation mésopotamienne. Éditions Robert Laffont (2001) pp. 545-546.
Kilmer, A., ‘The Cult Song with Music from Ancient Ugarit: Another Interpretation.’ Revue d’Assyriologie et d’Archéologie Orientale. Vol. 68. Éditions Presses Universitaires de France (1974) pp. 69-82.
Kilmer, A., ‘The Discovery of an Ancient Mesopotamian Theory of Music.’ Proceedings of the American Philosophical Society. Vol. 115. The American Philosophical Society (1971) pp. 131-149.
Kilmer, A., ‘Musik. A. I. In Mesopotamien.’ Reallexikon der Assyriologie und Vorderasiatischen Archäologie. Vol. 8. De Gruyter (1997) pp. 463-482.
Kilmer, A.; Crocker, R.L.; Brown, R.R., Sounds from Silence: Recent Discoveries in Ancient Near Eastern Music. Bit Enky Publications (1976).
Koitabashi, M., ‘Music in the Texts from Ugarit.’ Ugarit-Forschungen. Vol. 30. Ugarit-Verlag, Münster. Hubert & Co (1996) pp. 363-396.
rispijn, T.J.H., ‘Beitrage zur altorientalischen Musikforschung 1. Âulgi und die Musik.’ Akkadica. Vol. 70. Assyriological Center Georges Dossin (1990) pp. 1-27.
Lawergren, B., ‘Acoustics and Evolution of Arched Harps.’ The Galpin Society Journal. Vol. 34. Published by the Galpin Society (Oxford 1981) pp. 110-129.
Lawler, A., ‘Murder in Mesopotamia.’ Science. Vol. 317/5842. Published by AAAS (2007) pp. 1164-1165.
Lupton, C.J.; Huston, J.E.; Craddock, B.F.; Pfeiffer, F.A.; Polk, W.L., ‘Comparison of three systems for concurrent production of lamb meat and wool.’ Small Ruminant Research. Vol. 62/2. Official Journal of the International Goat Association. Published by Elsevier (2007) pp. 133-140.
Malamat, A., ‘Musicians from Hazor and Mari.’ Semitic and Assyriological studies presented to Pelio Fronzaroli. Harrassowitz Verlag (2003) pp. 355-357.
Marcetteau, M., ‘La musique au Proche-Orient ancien : l’approche des musicologues.’ Dossiers d’Archéologie. N° 310. Éditions Faton (2006) pp. 4-5.
Marcetteau, M., ‘A Queen’s Orchestra at the Court of Mari : New Perspectives on the Archaic Instrumentarium in the Third Millenium.’ ICONEA Proceedings 2008 (2008) pp. 67-73.
McMahon, A., ‘Report on the Excavations at Tell Brak, 2008.’ Newsletter of the British Institute for Study of Iraq. Vol. 22. Published by the British Institute for Study in Irak (2008) pp. 6-12.
McMahon, A., ‘Tell Brak 2007.’ Final Report. Published online (2007) pp. 1-7.
Menze, B.H.; Ur, J.A., ‘Mapping patterne of long-term settlement in Northern Mesopotamia at a large scale.’ Proceeding of the National Academy of Sciences – PNAS. Early Publication. Published by the National Academy of Sciences of the United States of America (2011) pp. 1-10.
Muldma, M., ‘La musique comme langue de communication dans le dialogue des cultures.’ Synergies – Pays Riverains de la Baltique. N° 6. Revue de Gerfint (2009) pp. 249-262.
Muldma, M., ‘Music as a Meeting Point of Cultures’. Education in Multicultural Environment, Dialogue of Cultures – Possibility or Inevitability ? Tome II. Tallinn University (2009) pp. 287-299.
Nemet-Nejat, K.R., ‘Music.’ Daily life in Ancient Mesopotamia. Published by Greenwood Press (1998) pp. 167-170.
Oates, J., ‘Archaeology in Mesopotamia: Digging Deeper at Tell Brak.’ Albert Reckitt Archaeological Lecture. Published by the British Academy (2004).
Oates, J.; Oates, D., ‘The Reattribution of Middle Uruk Materials at Brak.’ Leaving No Stones Unturned, Essays on the Ancient Near East and Egypt in Honor of Donald P. Hansen. Published by Eisenbrauns (2002) pp. 145-154.
Oates, J.; Oates, D., ‘The Role of Exchange Relations in the Origins of Mesopotamian Civilization.’ Explaining social change: studies in honour of Colin Renfrew. Published by the McDonald Institute for Archaeological Research, University of Cambridge (2004) pp. 177-192.
Oates, J.; Oates, D., ‘An Open Gate: Cities of the Fourth Millennium BC (Tell Brak 1997).’ Cambridge Archaeological Journal. N° 7. Published by The Cambridge University Press (1997) pp. 287-297.
Osses Adams, L., ‘Sumerian Harp from Ur.’ The American Harp Journal. Vol. 19/ 2. Published by the American Harp Society (2003) pp. 9-13.
Rebatet, L., Les Orchestres de Sumer. Une histoire de la musique. Éditions Robert Laffont (1973) pp. 17-29.
Risset, J.-C., ‘La musique et les sons ont-ils une forme ?’ La Recherche. Vol. 305. Éditions Sophia Publications (1998) pp. 98-102.
Rival, M., ‘Les instruments de musique au 4e millénaire.’ Les grandes inventions. Éditions Larousse (1994) pp. 40-41.
Ruwet, N., ‘Musicologie et linguistique.’ Étude international sur les tendances principales de la recherche dans les sciences de l’homme. Unesco/SS/41/3.244.1/h/24 (1966).
Schaeffner, A., ‘Travail et jeu.’ Origine des instruments de musique. Éditions EHESS (1994) pp. 95-108.
Seidel, H., ‘Musik und Religion. Altes und Neues Testament.’ Theologische Realenzyklopädie. Vol. 23. Éditions de Gruyter (1994) pp. 441-446.
Selz, G.J., ‘The Holy Drum, the Spear, and the Harp: Towards an Understanding of the Problems of Deification in Third Millennium Mesopotamia.’ Sumerian Gods and Their Representations. Published by STYX Publications (1997) pp. 167-213.
Shehata, D., ‘Les instruments de musique au Proche-Orient Ancien.’ Dossiers d’Archéologie. N° 310. Éditions Faton (2006) pp. 16-22.
Sloboda, J.A., ‘The Musical Mind, The Cognitive Psychology of Music.’ Oxford Psychology Series. N° 5. Published by Clarendon Press (1994).
Soltysiak, A., ‘Preliminary Report on Human Remains from Tell Majnuna (spring 2007).’ Excavations at Tell Brak 2006-2007. Published by the British Institute for Study in Irak (2007) pp. 161-163.
Soltysiak, A., ‘Short Fieldwork Report: Tell Majnuna (Syria), Season 2006.’ Bioarchaeology of the Near East. Vol. 2. Published online on www.anthropology.uw.edu.pl (2008) pp. 77-94.
Spycket, A., ‘La musique instrumentale mésopotamienne.’ Journal des savants. Vol. 3. Publication de l’Académie des Inscriptions et Belles Lettres. Diffusion De Boccard (1972) pp. 158-162. Tillmann, B., ‘La musique, un langage universel ?’ Pour la Science. N° 373. Éditions Belin (2008) pp. 124-138.
Valette, C.; Cuesta, C., Mécanique de la corde vibrante. Éditions Hermès (1993).
Wilson, S., ‘The effect of music on perceived atmosphere and purchase intentions in a restaurant.’ Psychology of Music. Vol. 31/1. Published by Sage Publications (2003) pp. 93-112.
BIBLIOGRAPHIE DU SYSTÈME AUDITIF
Alexander C.; Meyer, A.C. et al.,‘Tuning of synapse number, structure and function in the cochlea.’ Nature Neuroscience. Vol. 12/4. Nature Publishing Group (2009) pp. 444-453.
Arnold, W.; Anniko, M., ‘Supporting and membrane structures of human outer hair cells: evidence for an isometric contraction.’ ORL – Journal for Oto-Rhino-Laryngology and its related specialties. Vol. 51/6. Published by Karger (1989) pp. 339-353.
Ashmore, J., ‘Cochlear Outer Hair Cell Motility.’ The American Physiological Society. Vol. 88. Published by Wiley & Sons (2008).
Ashmore, J. et al., ‘The remarkable cochlear amplifier.’ Hearing Research. Vol. 266. Published by Elsevier (2010) pp. 1-17.
Bear, M.F.; Connors, B.W.; Paradiso, M.A.; Nieoullon, A., ‘Physiologie de la cochlée.’ Neurosciences. À la découverte du cerveau. 2e édition. Éditions Pradel (2002) pp. 370-389. Bremond, G. A., ‘Aspect général du labyrinthe et de son contenu conjonctif.’ L’oreille dans le temporal. Anatomie descriptive, topographique et systématisation. Éditions Solal (1994) pp. 14-44.
Breuskin, I.; Bodson, M.; Thelen, N.; Thiry, M.; Nguyen, L.; Belachew, S.; Lefebvre, P.P.; Malgrange, B., ‘Strategies to regenerate hair cells: Identification of progenitors and critical genes.’ The Journal of Hearing Research. Vol. 236/1-2. Published by Elsevier (2007) pp. 1-10.
Brownell, W.E., ‘Outer Hair Cell Electromotility and Otoacoustic Emissions.’ Ear Hear. Vol. 11/2. Published by Williams & Wilkins (1990) pp. 82-92.
Brownell, W.E.; Spector, A. A.; Raphael, R. M.; Popel, A. S., ‘Micro and nanomechanics of the cochlear outer hair cell.’ Annual Review Biomedical Engineering. Vol. 3. Published by Annual Reviews (2001) pp. 169-194.
Buser, P.; Imbert, M., ‘General structure of the sounds of musical instruments.’ Audition. The Physiological Psychology of Hearing. Vol.3.Massachusetts Institute of Technology (1992) pp. 95-111.
Cai, H.; Shoelson, B.; Chadwick, R.S., ‘Evidence of Tectorial Membrane Radial Motion in a Propagating Mode of a Complex Cochlear Model.’ PNAS (Proceedings of the National Academy of Sciences). Vol. 1/16. Published by the National Academy of Sciences of the United States of America (2004) pp. 6243-6248.
Corey, D. P.; Hudspeth, A. J., ‘Ionic basis of the receptor potential in a vertebrate hair cell.’ Nature. Vol. 281/5733. Published by Macmillan Journals Ltd (1979) pp. 675-677.
Dallos, P., ‘The Active Cochlea.’ The Journal of Neuroscience. Vol. 12/12. Published by The Society for Neuroscience (Washington 1992) pp. 4575-4585.
DeRosier,; D. J. Tilney, L. G., ‘The Structure of the Cuticular Plate, an In Vivo Actin Gel.’ The Journal of Cell Biology. Volume 109/6. Published by the Rockefeller University Press (1989) pp. 2853-2867.
Dulguerov, P.; Brownell, W. E., ‘Physiologie cochléaire.’ Précis d’audiophonologie et de déglutition. L’oreille et les voies de l’audition. Vol. 1. Éditions Solal (2005) pp. 57-79.
Eddine, C.A.; Williams, M.; Ayache, D., ‘Radio-anatomie utile de l’oreille.’ Journal of Radiology. Vol. 87. Éditions Elsevier Masson (2006) pp. 1728-1742.
Edeline, J.-M., ‘Anatomie et physiologie des voies auditives centrales : le système thalamo-cortical – L’oreille et les voies de l’audition’. Précis d’audiophologie et de déglutition. Tome I. Éditions Solal (2005) pp. 97-110.
Etournay, R.; Lepelletier, L.; Boutet de Monvel, J.; Michel, V.; Cayet, N.; Leibovici, M.; Weil, D.; Foucher, I.; Hardelin, J.-P.; Petit, C., ‘Cochlear outer hair cells indergo an apical circumference remodeling constrained by the hair bundle shape.’ Development. Vol. 137. Published by The Company of Biologists Ltd (2010) pp. 1373-1383.
Fettiplace, R.; Hackney, C.M., ‘The sensory and motor roles of auditory hair cells.’ Nature Reviews Neuroscience. Vol. 7. Published by Macmillan Journals Ltd (2006). pp. 19-29.
Forge. A.; Wright, T., ‘The molecular architecture of the inner ear.’ British Medical Bulletin. Vol. 63. Published by The British Council (2002) pp. 5-24.
Fraisse, P., ‘L’anticipation des stimuli rythmiques. Vitesse d’établissement et précision de la synchronisation.’ L’année psychologique. Vol. 66/1. Éditions Armand Colin (1966) pp. 15-36.
Fridberger, A.; Boutet de Monvel, J.; Zheng, J.; Hu,N.; Zou, Y.; Ren, T.; Nuttall,A., ‘Organ of Corti Potentials and the Motion of the Basilar Membrane.’ The Journal of Neuroscience. Vol. 2/45. Published by Wiley & Sons (2004) pp. 10057-10063.
Fuchs, P.A., ‘Time and intensity coding at the hair cell’s ribbon synapse.’ Journal of Physiology. Vol. 566/1. Published by The Physiological Society (2005) pp. 7-12.
Furness, D.N.; Mahendrasingam, S.; Ohashi, M.; Fettiplace, R.; Hackney, C.M., ‘The Dimensions and Composition of Stereociliary Rootlets in Mammalian Cochlear Hair Cells: Comparison between High- and Low-Frequency Cells and Evidence for a Connection to the Lateral Membrane.’ The Journal of Neuroscience. Vol. 28/25. Published by the Society for Neuroscience (2008) pp. 6342-6353.
Ghaffari, R.; Aranyosi, A.J.; Freeman, D.M., ‘Longitudinaly propagating traveling waves of the mammalian tectorial membrane.’ PNAS (Proceedings of the National Academy of Sciences). Vol. 104/42. Published by the National Academy of Sciences of the United States of America (2007) pp. 16510-16515.
Ghaffari, R.; Aranyosi, A.J.; Richardson, G.P.; Freeman, D.M., ‘Tectorial membrane travelling waves underlie abnormal hearing in Tectb mutant mice.’ Nature Communications. Vol. 1/96. Published by Nature Publishing Group (2010) pp. 1-6.
Gummer, A.W.; Hemmert, W.; Zenner, H.P., ‘Resonant tectorial membrane motion in the inner ear: Its crucial role in frequency tuning.’ PNAS (Proceeding of the National Academy of Sciences). Neurobiology. Vol. 93. Published by the National Academy of Sciences of the United States of America (1996) pp. 8727-8732.
Hanaway, J.; Woosley, T.A.; Gado, M.H.; Roberts, M.P., ‘Voies de l’audition.’ Atlas du cerveau. Un guide visuel du système nerveux central humain. Éditions De Boeck Université (2001) pp. 206-207.
Helmholtz, H.L.F., On the Sensations of Tone as a Physiological basis for a Theory of Music. Third Edition. Published by Longmans, Green, and Co. Aberdeen University Press (1895) reprinted by permission of Dover Publication, Inc. (1954).
Hemmert, W.; Zenner, H-P.; Gummer, A. W., ‘Three-Dimensional Motion of the Organ of Corti.’ Biophysical Journal. Vol. 78. Published by Cell Press (2000) pp. 2285-2297.
Imbert, M., ‘L’oreille et l’audition.’ Traité du cerveau. Éditions Odile Jacob (2006) pp. 233-248.
Kamina, P., ‘Voies cochléaires.’ Anatomie clinique. Tome 5. Éditions Maloine (2008) pp. 78-81.
Lewald, J.; Karnath, H.-O., ‘Vestibular Influence on Human Auditory Space Perception.’ Journal of Neurophysiology. Vol. 84/2. Published by The American Physiological Society (2000) pp. 1107-1111.
Lim, D., ‘Cochlear anatomy related to cochlear micromechanics’. Journal of Acoustical Society of America. Vol. 67/5. Published by the Acoustical Society of America, Melville (1980) pp. 1686-1695.
Lim, K.M.; Steele, C.R., ‘A three-dimensional nonlinear active cochlear model analyzed by the WKB-numeric method.’ Hearing Research. Vol. 170. Published by Elsevier (2002) pp. 190-205.
Augusto, L.S.C.; Kulay, L.A.; Franco, E.S., ‘Audition and exhibition to toluene – a contribution for the theme.’ International Archives of Otorhinolaryngology. Vol. 16/2. Open Access Journal (2012) pp. 246-258.
Lukashkin, A.N.; Lukashkina, V.A.; Legan, P.K.; Richardson, G.P.; Russell, I.J., ‘Role of the Tectorial Membrane Revealed by Otoacoustic Emissions.’ Journal of Neurophysiology. Vol.91/1. Published by The American Physiological Society (2004) pp. 163-171.
Marieb, E.N., ‘Oreille : ouïe et équilibre.’ Anatomie et physiologie humaines. Adaptation de la 6e édition américaine par R. Lachaîne. Éditions Pearson Education France (2005) pp. 599-608.
Marieb, E.N.; Hoehn, K., ‘The Ear: Hearing and Balance.’ Human Anatomy and Physiology. Seventh Edition. Published by Pearson Education (2004) pp. 583-592.
Nam, J.H.; Fettiplace, R., ‘Force Transmission in the Organ of Corti Micromachine.’ Biophysical Journal. Vol. 98. Published by Cell Press (2010) pp. 2813-2821.
Nilsen, K.E.; Russell, I.J., ‘Timing of cochlear feedback: spatial and temporal representation of atone across the basilar membrane.’ Nature Reviews Neuroscience. Vol. 2/7. Published by Macmillan Journals Ltd (1999) pp. 642-648.
Pritchard, T. C.; Alloway, K.D., ‘Système auditif.’ Neurosciences médicales. Les bases neuroanatomiques et neurophysiologiques. Éditions De Boeck Université (2002) pp. 277-298.
Pritchard, T.C.; Alloway, K.D., ‘Auditory System.’ Medical Neuroscience. First Edition. Fence Creek Publishing (1999) pp. 229-247.
Purves, D. Augustine, G. J. Fitzpatrick, D. Katz, L. C. Lamantia, A.S. McNamara, J.O., ‘Le système auditif.’ Neurosciences. Traduction de la 3e édition américaine par Jean-Marie Coquery. Éditions De Boeck (2005) pp. 283-314.
Purves, D.; Augustine, G.J.; Fitzpatrick, D.; Hall, W.C.; Lamantia, A.-S.; McNamara, J.O., ‘The Auditory System.’ Neuroscience. Third Edition. Published by Sinaur Associates Inc. (2004) pp. 283-314.
Raphael, Y.; Altschuler, R.A., ‘Structure and innervation of the cochlea.’ Brain Research. Vol.60. Published by Elsevier (2003) pp. 397-422.
Ren, T., ‘Longitudinal pattern of basilar membrane vibration in the sensitive cochlea.’ Proceeding of the National Academy of Sciences. Vol. 99/26. Published by the National Academy of Sciences of the United States of America (2002) pp. 17101-17106.
Richardson, G.P.; Lukashkin, A.N.; Russell, I.J., ‘The tectorial membrane: One slice of a complex cochlear sandwich.’ Current Opinion in Otolaryngology & Head and Neck Surgery. Vol. 16/5. Published by Lippincott Williams & Wilkins (2008) pp. 458-464.
Rival, M., ‘Musique et mathématiques.’ Les grandes inventions. Éditions Larousse (1994) p. 40.
Safieddine, S.; El-Amraoui, A.; Petit, C., ‘The Auditory Hair Cell Ribbon Synapse: From Assembly to Function.’ Annual Review of Neuroscience. Vol. 35. Published by Annual Reviews (2012) pp. 509-528.
Santos-Sacchi, J., ‘Functional motor microdomains of the outer hair cell lateral membrane.’ Pflügers Archiv European Journal of Physiology. Vol. 445. Published by Springer (2002) pp. 331-336.
Shoelson, B.; Dimitriadis, E.K.; Cai,H.; Kachar, B.; Chadwick, R.S., ‘Evidence and Implications of Inhomogeneity in Tectorial Membrane Elasticity.’ Biophysical Journal. Vol. 87. Published by Rockefeller University Press (2004) pp. 2768-2777.
Stepanyan, R.; Belyantseva, I.A.; Griffith, A.J.; Friedman. T,B.; Frolenkov. G.I., ‘Auditory mechanotransduction in the absence of functional myosin-Xva.’ The Journal of Physiology. Vol. 576/3. Published by the Cambridge University Press (2006) pp. 801-808.
Temchin, A.N.; Recio-Spinoso, A.; Cai, H.; Ruggero, M.A., ‘Traveling Waves on the Organ of Corti of the Chinchilla Cochlea: Spatial Trajectories of Inner Hair Cell Depolarization Inferred from Responses of Auditory-Nerve Fibers.’ The Journal of Neuroscience. Vol. 32/31. Published by The Society for Neuroscience (2012) pp. 10522-10529.
Tilney, L.G.; Derosier, D.J.; Mulroy, M.J., ‘The Organization of Actin Filaments in the Stereocilia of Cochlear Hair Cells.’ The journal of Cell biology. Vol. 86/1. Published by the Rockefeller University Press (1980) pp. 244-259.
Tsuprun, V.; Santi, P., ‘Ultrastructural organization of proteoglycans and fibrillar matrix of the tectorial membrane.’ Hearing Research. Vol. 110/1-2. Published by Elsevier (1997) pp. 107-118.
Ulfendahl, M.; Flock, A., ‘Outer Hair Cells Provide Active Tuning in the Organ of Corti.’ Physiology. News in Physiological Sciences. Vol. 13. Published by Waverly Press (1998) pp. 107-111.
Verpy, E.; Leibovici, M.; Michalski, N.; Goodyear, R.; J. Houdon, C.; Weil, D.; Richardson, G. P.; Petit, C., ‘Stereocilin connects outer-hair-cell stereocilia to one another and to the tectorial membrane.’ The Journal of Comparative Neurology. Vol. 519/2. Published by the Wistar Institute of Anatomy and Biology (2011) pp. 194-210.
Vranceanu, F.; Perkins, G.A.; Terada, M.; Chidavaenzi, R.L.; Ellisman, M.H.; Lysakowski, A., ‘Striated organelle, a cytoskeletal structure positioned to modulate hair-cell transduction.’ PNAS (Proceeding of the National Academy of Sciences). Vol. 109/12. Published by the National Academy of Sciences of the United States of America (2012) pp. 4473-4478.
Yu, N.; Zhao, H-B., ‘Modulation of Outer Hair Cell Electromotility by Cochlear Supporting Cells and Gap Junctions.’ Plos One : www.plosone.org. Vol. 4/11. Published online (2009).
Zetes, D.E.; Steele, C.R., ‘Fluid-structure interaction of the sterocilia bundle in relation to mechanotransduction.’ Journal of Acoustical Society of America. Vol. 101/6. Published by the Acoustical Society of America (1997) pp. 3593-3601.
Zheng, J.; Shen, W.; He, D.Z.Z.; Long, K.B.; Madison, L.D.; Dallos, P., ‘Prestin is the motor protein of cochlear outer hair cells.’ Nature. Vol. 405. Nature Publishing Group (2000) pp. 149-155.
TABLE DES ILLUSTRATIONS
Illustration de la ‘page à la une’ : Infographie Girard, T. Arrière-plan d’après une photographie de Lenoir, M. Inserm. Promenade autour de la cochlée. Modifiée.
1) (à gauche) Girard, T. d’après Butterlin, P. 2003. (en haut à droite) Menze, B. & Ur, J. 2011. (en bas à droite) © McMahon, A. 2008. Report on the excavations at Tell Brak. Tell Brak Project. Courtoisie de l’auteur.
2) A) Amiet, P. 1980. La glyptique mésopotamienne archaïque. Courtoisie de l’auteur. B) Becker, A. 1993. C), D) et E) Oriental Institute University of Chicago. © Hirmer Verlag Munich. Gros plans : Lindemeyer, E. & Martin, L. 1993.
3) (à gauche) A) Delouzag, P.P. Kantor, H.J. 1972. B) Delougaz, P. et Kantor, H.J. 1971. Avec l’aimable autorisation de The Oriental Institute of the University of Chicago. (à droite) Amiet, P. 1980. La glyptique mésopotamienne archaïque. Courtoisie de l’auteur.
4) A) et B) Amiet, P. 1980. La glyptique mésopotamienne archaïque. Courtoisie de l’auteur. C), D), E) et F) CDLI n° P003174, n° P001443, n° P000887, n° P001401. © German Archaeological Institute (DAI). Berlin. Germany.
5) (à gauche) Girard, T. sous la direction de Dumbrill, R. (à droite) Girard, T. sous la direction de Dumbrill, R. 6) A) et B) Amiet, P. 1980. La glyptique mésopotamienne archaïque. Courtoisie de l’auteur. C) © Iraq Museum. Bagdad. D) © R.M.N. Photographie Galland, J.
7) (à gauche) A) Girard, T. D’après Marieb, E.N. Anatomie et physiologie humaines. 2005. P 604. Fig 15.29. B) Girard, T. Photographie de Welleschik, B. 2006. Creative Commons. (en haut à droite) Girard, T. D’après la photographie de Leblanc, A. 1998. Atlas des organes de l’audition et de l’équilibration. P 41. Fig 26 b. (en bas à droite) A) Girard, T. D’après Kamina, P. 1996. P 168. B) Photographie de la cochlée. Inserm. Pujol, R. Site Internet Promenade autour de la cochlée.
8) A) Girard, T. B) Girard, T. D’après un dessin publié sur le site Internet de l’Inserm, Promenade autour de la cochlée. C) Girard, T. D’après Kamina, P. 1996. Anatomie. Tête et cou. P 170. D) Girard, T. D’après Dulguerov, P. Brownell, M.E. 2005. Précis d’audiophonologie et de déglutition. P 57. Fig 4.1. Imbert, M. Buser, P. 1987. P 6. Fig 1.2. P 184. Fig 2.55.
9) (à gauche) A) Carrat, R. 2009. P 41. Fig 2.10. Netter, F. Myers, D. 1962. B) Girard, T. D’après une source non-connue. (en haut à droite) Dulguerov, P. Brownell, M.E. 2005. Précis d’audiophonologie et de déglutition. P 52. Fig 3.7. Photograph by Chevallier, J-M.; De Bonfils, P. Avec l’aimable autorisation des éditions De Boeck-Solal. (en bas à droite) A) Kros, C.J. Crawford, A.C. 1990. P 266. Fig 1. Courtoisie des auteurs. B) Frolenkov, G.I. 2006. Regulation of electromotility in the cochlear outer hair cell. P 44. Fig 1-C. Courtoisie de l’auteur. C) Leibovoci, M. 2006. Institut Pasteur. Courtoisie de l’auteur. D) Maltby, M. 2005. Occupational Audiometry. P 8. Fig 1.3. Courtoisie de l’auteur.
10) A) Girard, T. Leibovici, M. Sans, A. B) Girard, T. et Leibovici, M. C) Reconstitution par infographie de la membrane tectoriale. Girard, T. à partir d’une photographie de Lenoir, M. Inserm. Promenade autour de la cochlée. D) Girard, T. Leibovici, M. Sans, A.
11) (à gauche) A) « La Harpiste » dessin modifié © 2011 Swaze – site : www.swaze.fr. Courtoisie de l’auteur. B) Girard, T. Leibovici, M. (en haut à droite) Reconstitution par infographie. Girard, T. Dumbrill, R. (en bas à droite) Photographie à gauche : Parsa, A. et al. 2012. Deiters cells tread a narrow path – The Deiters cells-basilar membrane junction – P 14. Fig 2-C. Photographie à droite : Leibovici, M. 2012. Courtoisie des auteurs.
12) A) Girard, T. Dumbrill, R. Photographies Chadefaux, D. et al. 2010. Experimental study of the plucking of the concert harp. P 2. Fig 2. Courtoisie de l’auteur. B) Girard, T. Dumbrill, R. C) Girard, T. Leibovici, M. Photographies Frolenkov, G. et al. 1998. The Membrane-based Mechanism of Cell Motility in Cochlear Outer Hair Cells. P 1962. Fig 1. Fettiplace, R. Hackney, C.M. 2006. The sensory and motor roles of auditory hair cells. P 26. Fig 5. D) Girard, T. Leibovici, M. D’après Frolenkov, G. et al. 1998. Ashmore, J. 2008.
13) (à gauche) A) et B) Girard, T. Dumbrill, R. Ondulations 3D d’après Yang, J. et al. 2012. Interaction of highly nonlinear solitary waves with thin plates.P 1465. Fig 2. Photographie : Delalleau, A. et al. 2009. Un modèle hyperélastique à réorientation de fibres pour l’analyse des caractéristiques mécaniques de la peau. Fig 1-III. (en haut et en bas, à droite) Girard, T. Leibovici, M. Ondulations 3D d’après Yang, J. et al. 2012. Un modèle hyperélastique à réorientation de fibres pour l’analyse des caractéristiques mécaniques de la peau. P 1465. Fig 2. Photographie : Shoelson, B. 2004. Evidence and Implications of Inhomogeneity in Tectorial Membrane Elasticity. P 2772. Fig 8 © Elsevier. Schéma : Girard, T. d’après Meaud, J. Grosh, K. 2011. Coupling Active Hair Bundle Mechanics, Fast Adaptation, and Somatic Motility in a Cochlear Model. P 2579. Fig 1A.
14) A), B) et C) dessin et infographie Girard, T. D) Site Internet Voyage au centre de l’audition, Pujol, R. E) Ross, M. D. 1974. Tectorial Membrane of the Rat. Planche 3.14. F) Shoelson, B. et al. 2004. ‘Evidence and Implications of Inhomogeneity in Tectorial Membrane Elasticity.’ Biophysical Journal. Vol. 87. P 2769 Fig 2. © Elsevier. Avec l’aimable autorisation de l’éditeur.
15) (à gauche) A) Girard, T. Dumbrill, R. B) Girard, T. Leibovici, M. (en haut à droite) Girard, T. Dumbrill, R. (en bas à droite) Girard, T. Leibovici, M. D’après Safieddine, S. 2012. The Auditory Hair Cell Ribbon Synapse: From Assembly to Function. P 511. Fig 1-D.
16) Girard, T. Roi. P. (encadré en bas à droite) Harpe Arquée E. 2483. @ Cité de la musique.
© 2013. La Théorie Sensorielle.
